Diagnosis, Species Identification, and Genotyping
Preliminary diagnosis of cryptococcosis is made by direct microscopic examination of India ink preparations of cerebrospinal fluid samples or rapid staining (DiffQuik, Giemsa, etc.) of smears made from aspirates and nasal exudates (Kwon-Chung and Bennett 1992) (Fig.
12.6). In fresh material, India ink preparations often displayFig. 12.6 Photomicrograph of a DiffQuik®-stained smear from nasal exudate from a cat with sinonasal cryptococcosis. The negative-staining capsule is characteristic, as is the rare narrow-necked budding
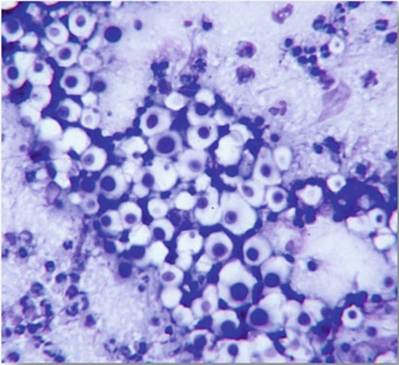
spherical vacuoles in Brownian motion (Mitchell and Perfect 1995) (Fig. 12.7). The capsules are generally spherical and may appear thick and vary in diameter.
If mycologically suitable and if suitable substrate is present in the media, culturing is the most suitable method for demonstrating the presence of pathogenic fungi (Vanessa et al. 2005). Cryptococcus may be cultured on routine mycological culture medium (Sabouraud glucose agar), on which it produces white, variably mucoid colonies when inoculated at 37 °C. Unlike other saprophytic species, C. neoformans produces laccase (phenol oxidase), which leads to the formation of melanin. Colonies turn brown to black and may thus be readily be identified (either C. gattii or C. neoformans); therefore, any medium, e.g., caffeic acid, Staib's medium, or tobacco agar medium, containing a substrate for phenol oxidase may serve as the primary isolation medium for C. neoformans and C. gattii. This is especially useful when sampling non-sterile sites (e.g., the nasal cavity) or the environment, where other fungi are often present. Both species may be recognized by color differentiation when grown on Christensen urea agar and L-canavanine-glycine-bromothymol blue (CGB) agar.
Despite the incidence of false-positive and false-negative results, CGB medium is most widely used for differentiating between C. gattii and C. neoformans (McTaggart et al. 2011; Klein et al. 2009).Histopathology, which may be used to investigate the presence of the fungus in tissue sections via corresponding tissue reactions, provides useful insights (Chander 2009). Cryptococcal cells may be easily detected in tissue sections of the lungs, brain, and liver using specific stains such as Gomori's methenamine silver nitrate, periodic acid-Schiff and Mayer's mucicarmine (Fig. 12.8). However, in
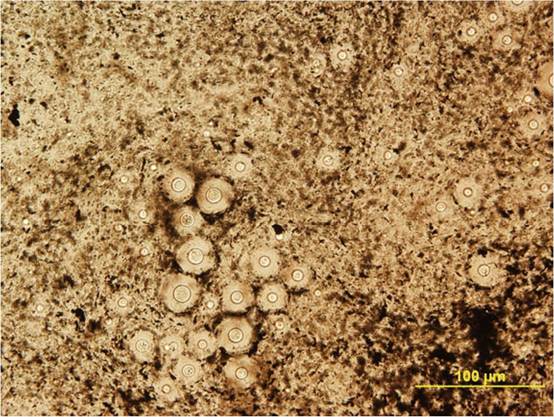
Fig. 12.7 Indian ink preparation of cerebrospinal fluid (CSF) from a dog with cryptococcal meningoencephalitis. Note the negatively stained capsules thrown into relief by the dark carbon material in the Indian ink stain
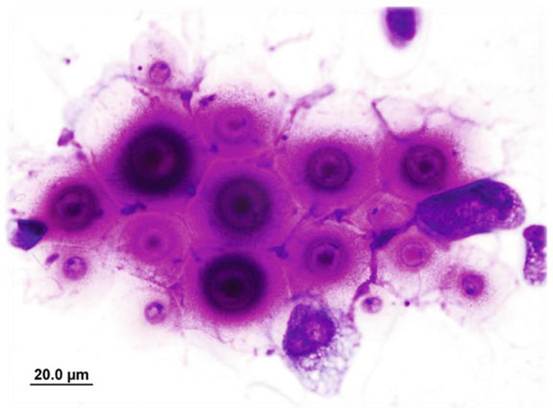
Fig. 12.8 Photomicrograph of histology from a dog with widely disseminated cryptococcosis. The mucicarmine stains the capsule bright pink. Image courtesy of Alan Kessel
hematoxylin-eosin-stained sections, tissue reactions are poorly demonstrated, and diagnosis therefore depends on the observation of cryptococcal capsules within the tissues.
Some physiological features of C. neoformans, such as urease production (in the presence of urea), have been used as the basis of differentiations between isolates (Singh et al. 2013). The presence of this enzyme can be determined and semiquantitated by the rapid urease test and using Christensen’s agar medium. Similarly, in yeast carbon base medium, C. gattii assimilates D-proline, whereas C. neoformans usually fails to do so (Dufait et al. 1987). Wickerham and Burton (1948) have reemphasized the role of carbon assimilation tests in the taxonomy of Cryptococcus. In most clinical laboratories worldwide, this method has been replaced by several commercial kits (Kwon-Chung and Bennett 1992).
The physiological tests commonly used for identification are fermentation of seven to eight carbohydrates, growth on various carbon and nitrogen sources, determination of growth requirements, growth on media with a high content of sugar, and testing for urea hydrolysis (Rij 1984). There is no correlation between the growth of yeast on a particular carbohydrate and fermentation of the same carbohydrate by that yeast. Yeasts vary in their ability to ferment sugars, which may be measured by the production of CO2. C. neoformans and C. gattii assimilate sucrose and maltose but are usually lactose negative (Kwon-Chung and Bennett 1992).Carbon assimilation tests determine the ability of yeast to grow aerobically on a particular carbon compound (Rij 1984). These tests may be performed either on solid or liquid media (Wickerham 1951). Yeast nitrogen base medium is most commonly used in carbon assimilation tests. Similarly, yeasts are additionally capable of utilizing a wide variety of nitrogen sources. Various nitrogen compounds, such as nitrate, nitrite, ethylamine hydrochloride imidazole, ammonium sulfate, glucosamine, creatine, and creatinine, are also tested by carbon assimilation tests.
The rapid detection of the presence of yeast in clinical specimens has also been performed by several molecular methods. Numerous molecular approaches have shown great potential in the field of mycology diagnostics during the last decade. Molecular methods based on the analysis of yeast nucleic acids were first used for the investigation of total nuclear DNA and the determination of the existence of co-specific relationships between morphologically and physiologically different strains (Alcoba-Florez et al. 2007). Historically, different isolates of the same species were distinguished by biotyping or serotyping. Currently, various molecular techniques have superseded older methodologies, including PCR-fingerprinting, restriction fragment length polymorphism (RFLP) analysis, amplified fragment length polymorphism (AFLP) analysis, multilocus sequence typing (MLST), and microsatellite length polymorphism (MLP) using short tandem repeat (STR) markers.
These methods enable the researcher to determine whether isolates belong to a single strain (biotype). In addition, next-generation sequencing of the whole fungal genome is becoming the most appropriate method for fungi with no MLP or MLST schemes available (Loftus et al. 2005; Alanio et al. 2017).Karyotyping was one of the first molecular methods employed for the identification of Cryptococcus (Perfect et al. 1989). C. neoformans and C. gattii have been distinguished in terms of number and lengths of chromosomes by contour-clamped homogenous field gel electrophoresis and orthogonal-field-alternation gel electrophoresis (Polacheck and Lebens 1989; Boekhout et al. 1997; Hagen 2011).
Highly sensitive and rapid PCR amplification possess advantages over various molecular genotyping methods. Several PCR protocols have been used for the detection and identification of fungal species (Chang et al. 2001). However, methods of DNA extraction and purification have not been standardized to date (Alcoba- Florez et al. 2007). Various PCR-based approaches, such as PCR with specific primers at the genus or species level, nested PCR, real-time PCR, multiplex PCR, PCR and posterior restriction analysis of amplicons, and PCR followed by sequencing, have been applied (Ahmad et al. 2002; Alcoba-Florez et al. 2005, 2007). RFLP fingerprinting utilizes the rDNA locus to fingerprint the members of genus Cryptococcus, although C. neoformans could not be differentiated from C. gattii following the digestion of PCR products with four restriction enzymes (Vilgalys and Hester 1990; Hagen 2011). Researchers tested several hybridization probes such as the M13 phage core sequence (GACA)4 and (GTG)5, which are based on hypervariable, repetitive, and highly conserved DNA sequences/regions (Meyer et al. 1993; Meyer and Mitchel 1995). Hypervariable regions differing in sequences and/or length permit the identification of Cryptococcus at the species level. Overall, four genotypes were designated: VNI and VNII for C.
neoformans var. grubii (serotype A), VNIII for hybrid (serotype AD), and VNIV for C. neoformans var. neoformans (serotype D) using random amplification of polymorphic DNA (RAPD) (Meyer et al. 1993). Subsequently, four genotypes were also introduced for C. gattii, namely, VGI, VGII, VGIII, and VGIV (Sorrell et al. 1996). The species C. neoformans was subsequently investigated by the analysis of the intergenic spacer (IGS) region (rDNA sequence) and internal transcribed spacer (ITS), a phylogenetically important region of DNA (Diaz and Fell 2000; Diaz et al. 2005; Imai et al. 2000). IGS revealed significant differences between and within the varieties of C. neoformans and further indicated a similar grouping of C. neoformans isolates (Katsu et al. 2004; Hagen 2011). Furthermore, researchers combined the sequences of IGS, ITS, and topoisomerase and applied MLST (Sugita et al. 2001). Subsequently, a consensus MLST scheme was proposed (Meyer et al. 2009), comprising of seven unlinked nuclear loci CAP59, GPDi, IGSi, LACi, PLBi, SODi, and URA5, with optional discriminatory power. Recently, multiple multilocus microsatellite typing (MLMT) schemes have been developed for the two sibling species of Cryptococcus; these are more reliable and cost-effective than MLST (Karaoglu et al. 2008; Illnait-Zaragozi et al. 2010).A relatively novel method using the matrix-assisted laser desorption/ionization- time-of-flight (MALDI-TOF) mass spectrometry (MS)-based strategy has shown promising results to discriminate major molecular types within the C. neoformans and C. gattii species complexes obtained from human (Firacative et al. 20i2) and veterinary sources (Danesi et al. 20i4b). Currently, two MALDI-TOF MS systems are approved by the US Food and Drug Administration for the identification of yeasts: the Bruker Biotyper (Bruker Daltonics, Bremen, Germany) and the Vitek MS (bioMerieux, Marcy l'Etoile, France) systems. The Bruker instrument provides the MALDI Biotyper (software and database), and bioMerieux includes the Vitek MS and the SARAMIS (AnagnosTec, Germany) databases, referred to as the Vitek MS IVD system.
Biotyper software generates log score values ranging from 0 to 3, with a value of ≥2 indicating species-level identification; scores between i.7 and i.99, genus-level identification; and scores below i.7 unreliable identification. Meanwhile, using Vitek MS software, a confidence score value of ≥60% is recommended for species-level identification (Posteraro et al. 20i2). The species coverage of MALDI-TOF MS reference databases represents the Achilles' heel of each MALDI-TOF MS system. It is therefore recommended for users to create and supplement their own libraries of reference mass spectra by including locally important strains or species/strains not represented or not sufficiently represented in the commercial libraries (Posteraro et al. 20i3).Diagnosis of animal cryptococcosis can also be achieved by using serological assays that detect the capsular glucuronoxylomannan polysaccharide of Cryptococcus species in body fluids. Serum is the specimen of choice for antigen testing, although cerebrospinal fluid and urine can also be used (Boulware et al. 20i4; Jarvis et al. 20ii). Multiple assays have been developed and approved by the FDA for the detection of the capsular antigen of Cryptococcus species, including a Cryptococcus antigen (CrAg) latex agglutination system (CALAS) (Meridian Bioscience, Cincinnati, OH, USA) and a CrAg lateral flow assay (LFA;IMMY, Norman, OK, USA) (Temstet et al. i992; Binnicker et al. 20i2). In veterinary medicine, the CALAS assays have been most widely used to date. Antigen titers are frequently high to extremely high (>i:65,536) in cats, but even a titer of i:2 can indicate cryptococcal infection (Sykes and Malik 20i4). Titers typically decline with successful treatment, although the drop in titer generally lags behind the clinical improvement. False-negative results in cats appear to be very rare (sensitivity >95%) but can occur in cases of very localized infections; therefore, a negative titer does not completely rule out cryptococcosis (Belluco et al. 2008). The causes of false-positive test results in these situations remain unclear. Before treatment is initiated, the diagnosis of cryptococcosis should be confirmed using other diagnostic tests (cytology and culture) in cats with weakly positive antigen titers. The CALAS test is expensive and requires extra technological input; thus, the method is not practical in many developing countries, where cryptococcosis is endemic or enzootic. In contrast, the novel point-of-care CrAg LFA offers several advantages over the CALAS, including enhanced sensitivity for the detection of CrAg and a rapid turnaround time of approximately 15 min. In addition, the assay does not require pronase pretreatment of serum samples (Jarvis et al. 2011; McMullan et al. 2012).
12.9