Epidemiology of Human and Animal Coccidioidomycosis
In the 1940s, one of the first investigations into the source of infection for a localized outbreak of CM was conducted (Davis et al. 1942). In April 1940 on a field trip to San Benito County, California, several Stanford students and faculty spent 2 days collection specimens and camping.
Ten days after return to campus, a student was seen at the health center with fever, chest congestion, and malaise. Five more students from the same field trip reported similar illness within the week. Although the other students recovered, the first student that had become ill was diagnosed with coccidioidomycosis. Eventually, the group exposure was determined to the result of students digging a rattlesnake out of ground squirrel burrow. Coccidioides immitis was cultured out of the soils that were collected at the presumed exposure site.Phylogenomic analyses reveal that C. immitis and C. posadasii speciated between five and ten million years ago (Engelthaler et al. 2015; Sharpton et al. 2009). The genus Coccidioides is possibly as old as 40-50 million years (Bowman et al. 1992; Fisher et al. 2002). Many mammalian orders rapidly diversified during the early Cenozoic, and the appearance of rodent fossils in North America corresponds to the proposed emergence of the Coccidioides genus (Fisher et al. 2002; Tapaltsyan et al. 2015; Saarinen et al. 2014). The emergence of both the Sierra Madres and Rocky Mountains in North America corresponds with divergence among early North American mammals (Saarinen et al. 2014; Bally and Palmer 1989). As South America was a separate continent until approximately three million years ago, recent introduction of Coccidioides posadasii to that region, and the origin of Coccidioides in the Sonoran Desert, is supported by both genomics and geological records (Engelthaler et al. 2015).
4.4.1 Climate
Climate clearly influences the incidence of CM and presence of the organism in the environment.
Early investigations into the role of climate at Williams Air Force Base in Maricopa County during WW2 showed two seasonal increases in infection rates after winter and summer precipitation (Hugenholtz 1957). A similar trend among Arizona residents has been observed more recently (Tamerius and Comrie 2011; Kolivras and Comrie 2003). In Kern County California, only a single increase in infection rate occurs after the winter rainy season (Talamantes et al. 2007). Climatic factors that might affect rates in other endemic area, specifically in Mexico and South America, are unknown at this time (Vargas-Gastelum et al. 2015; Baptista- Rosas et al. 2007, 2012). The recent autochthonous infections in Washington state raise concerns, as there is not a consensus on the effect climate change will have on the spread of the organism into new areas (Litvintseva et al. 2015). The fungus has been found sporadically at Dinosaur National Monument in northern Utah, which indicates suitable habitat can be found in the Great Basin Desert (Johnson et al. 2014; Fisher et al. 2007; Petersen et al. 2004). Climate change predicts less frequent but more intense precipitation, and higher mean temperatures in the western USA, which may expand the endemic region.4.4.2 Environmental Niche
Current understanding of the distribution of the fungus in the environment is based on extensive skin testing using a delayed-type hypersensitive reaction to various Coccidioides antigens (Fiese 1958). Using either spherulin (antigen derived from parasitic growth phase) or coccidioidin (antigens derived from saprobic growth phase), researchers mapped highest disease prevalence in the southwestern USA (Pappagianis 1988; Ajello 1971). Long-term Arizona residents in Maricopa County (Phoenix), Pima County (Tucson), and Pinal County (Casa Grande, Florence) had over 70% positive skin test rates, when compared to surrounding counties with only 10-40% reactivity rates (Maddy 1957, 1958b; Palmer et al. 1957; Edwards and Palmer 1957).
Similar testing completed in California shows that Kern County (Bakersfield), Tulare County (Visalia), and Kings County (Hanford) residents had 50-70% skin test positive rates, while in surrounding counties positive skin tests dropped to 10% (Edwards and Palmer 1957). Additionally, in Mexico, Central America, and South America, similar distributions of positive skin tests have been found (Campins 1970; Mayorga and Espinoza 1970). However, in sparsely populated regions, this approach may not reflect the distribution of Coccidioides in the environment. Moreover, human migration over large and small spatial scales confounds fine spatial-scale analyses.Several groups have worked to understand the ecology of Coccidioides (Whiston and Taylor 2014; Barker et al. 2012; Baptista-Rosas et al. 2007; Lacy and Swatek 1974; Swatek and Omieczynski 1970; Teel et al. 1970; Elconin et al. 1964). Defining factors that determine the presence of Coccidioides, as well as distribution of the fungus in the soil at local sites, has been investigated (Litvintseva et al. 2015; Johnson et al. 2014; Barker et al. 2012; Baptista-Rosas et al. 2007, 2012; Kolivras and Comrie 2003; Greene et al. 2000; Lacy and Swatek 1974; Swatek 1970; Swatek and Omieczynski 1970; Egeberg and Ely 1956). Environmental isolates of Coccidioides are usually obtained via inoculation of soil extracts in a susceptible rodent model (Davis et al. 1942; Levine and Winn 1964). The distribution of the fungus in the environment has been determined to be sporadic and highly localized, on the order of a square meter or less in area (Maddy 1958b, 1965). Additionally, trapping of rodents at the positive sites was conducted, with low overall levels of infectivity (Emmons 1942, 1943; Emmons and Ashburn 1942).
Defining key factors that explain presence of Coccidioides in the environment remains an elusive problem. Associations with saline and alkaline soils are the pattern in California (Plunkett et al. 1963; Egeberg et al. 1964; Elconin et al.
1964). However, in Arizona sandy and porous soil along with rodent burrows appears to be the strongest association (Barker et al. 2012; Emmons 1942; Maddy 1959, 1965; Smith 1971; Swatek 1970). However, caution is warranted when attempting to generalize the results, due to few studies and variable design of collection. Overall, most soil samples tested are negative (ranges from 99 to 80%), and completely randomized sampling approaches have resulted in predominately negative results (Lacy and Swatek 1974; Greene et al. 2000; Lauer et al. 2012;Barker et al. 2012; Baptista-Rosas et al. 2012). Complex relationships among microbial organisms that share the same habitats have been investigated (Egeberg et al. 1964; Orr 1968). Direct plating from soil often results in overgrowth by more rapidly growing fungi under laboratory growth conditions (Swatek and Omieczynski 1970; Greene et al. 2000; Barker et al. 2012). However, Coccidioides is competitive under certain circumstances and may persist for decades the same location (Barker et al. 2012; Greene et al. 2000).
Distinct population and species boundaries, both within and among each species, are still unclear due to evidence of hybridization and introgression and that both species have been recovered among patients in southern California and northern Mexico (Neafsey et al. 2010; Fisher et al. 2001, 2002; Canteros et al. 2015; Johnson et al. 2014; Litvintseva et al. 2015; Lauer et al. 2012). MLST analysis of over 600 clinical and environmental isolates reveals population structure within Arizona and that clinical isolates are distinct from environmental isolates (Teixeira and Barker 2016).
Additionally, techniques to detect the fungus in the environment are being developed, which will help to understand and define the environmental niche of Coccidioides. New work on air sampling has provided a needed tool to monitor seasonal fluctuations (Chow et al. 2016). Extracting DNA from soil and dust has become much more common and molecular methods to detect the organism more robust (Johnson et al.
2014; Lauer et al. 2012; Baptista-Rosas et al. 2012; Litvintseva et al. 2015). Although detecting Coccidioides DNA in soil does not prove the presence of infectious arthroconidia, it is a method for screening a large number of soil samples, which would be necessary to model the environmental niche of the fungus.4.4.3 Substrate Preferences
Comparative genomic studies have revealed functional differences associated with pathogenic, saprobic, or commensal lifestyles. Coccidioides species can digest keratin and other animal products and appear to have lost many genes associated with plant-derived carbon sources (Whiston and Taylor 2014, 2015; Sharpton et al. 2008). This suggests that this genus has specialized on animal-derived nutrients. This could be either acquired from tissue digestion during the parasitic phase in vivo or could be dead, decaying, or other sources of keratin such as skin, feathers, and hair (Lange et al. 2016; Lopes et al. 2008). It is likely that the primary nutritional mode is closely associated with animal-derived sources and may have led to the evolution of the parasitic lifestyle (Whiston and Taylor 2014; Sharpton et al. 2009).
Specifically, comparisons between Onygenales and Eurotiales (sister orders) show a reduction of cellulose-binding domain containing proteins, tannases, cellulases, cutinases, melibiases, pectin lyase, and pectin esterases among the Onygenales (Sharpton et al. 2009). These are all classes of genes associated with plant-derived nutritional sources. Two families appear to be expanded only in Coccidioides, and in particular the M35 class of deuterolysin metalloproteases, which contains at least one known virulence factor Mep1 (Hung et al. 2005; Whiston and Taylor 2015). This family of proteases has a preferred substrate of histones and protamines, which are arginine-rich molecules (Doi et al. 2003). This class is also under positive selection, supporting the idea that these genes are associated with the evolution of Coccidioides (Li et al.
2012). It is suggested that keratin degradation is associated with M35 class of metalloproteases, and thus Coccidioides may be associated preferentially with animal-derived nutritional sources, rather than plant- derived nutrients. This remains to be experimentally proven, and it is possible that the role of M35 deuterolysin metalloproteases in Coccidioides has diverged into new, and as yet unexplored, functions.Other animal-associated nutrition sources include dung and frass. Fecal samples taken from lizards (Uta Stansburiana, Gerrhonotus spp., Sceloporus occidentalis, Crotaphytus wislizeni, Cnemidophorus tigris), skunks (Spilogale gracilis), blacktailed deer (Dama hemionus), goats (Capra spp.), sheep (Ovis spp.), and burros (Equus asinus) near positive soil locations were subjected to culture to attempt to recover Coccidioides; however, none grew the organism (Swatek et al. 1967). Coccidioides has been cultured from bat guano in a single report (Krutzsch and Watson 1978).
4.4.4 Range Expansion
Coccidioides spp. are found in arid or semi-arid regions throughout the Americas but are thought to be at highest prevalence in southern Arizona (Fig. 4.2) (Fisher et al. 2007). C. immitis is found in central and southern California (Fisher et al. 2001). The range extends into northern Mexico, and recent work has found C. immitis in Yakima and Benton counties of Washington, at Dinosaur National Monument in Utah, and from a patient in Colombia with no travel history (Marsden-Haug et al. 2014; Litvintseva et al. 2015; Johnson et al. 2014; Canteros et al. 2015). Coccidioides posadasii is found in Arizona, Nevada, Utah, New Mexico, Texas, and throughout Mexico, with dispersed populations in Central and South America (Whiston and Taylor 2014; Duarte-Escalante et al. 2013; Brilhante et al. 2013; Campins 1970). Hybrid strains indicate that the two species coexist in nature (Neafsey et al. 2010). To determine if this is a recent or ancient phenomenon, environmental sampling is needed to accurately assess the prevalence of both species, and hybrid offspring, at a given location.
Direct isolations from soil throughout the range of both species in North and South America will clarify population structure and species boundaries. Clinical isolates are still needed to track emergence of any virulent strains or new point source outbreaks (Litvintseva et al. 2015). With greater surveillance and awareness, it is predicted that potential habitat for and cases of Coccidioides may be found throughout the western USA (Baddley et al. 2011). In fact, recent reports of cases in
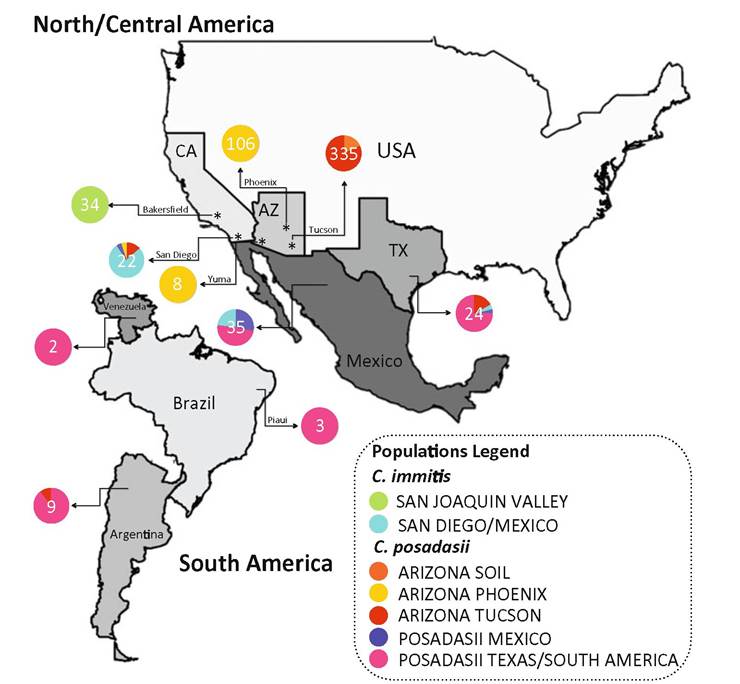
Fig. 4.2 Distribution and prevalence of Coccidioides spp. in arid or semi-arid regions throughout the America
Missouri northeast of the endemic area are concerning (Turabelidze et al. 2015). Although many of these cases could be the result of travel to endemic areas, the possibility must be considered that these are locally acquired infections as a result of drier soil and dust storms, fomites, or even unrecognized small foci of growth of the fungus (Hage et al. 2012; Stagliano et al. 2007; Desai et al. 2001).
4.5