Gross anatomy of the intestinal tract
The intestinal tract is essentially a muscular tube, with a varying diameter and mucosal structure in the different sections of the tract. The variations in diameter, mucosal structure and function relate to the differing physiological role of each area; for instance, the stomach is readily distensible, while the small intestine is much less distensible.
The intestinal wall is divided into four discrete layers. The four layers are visible on cut sections of the intestine and can be readily distinguished by differences in echogenicity during ultrasound examination of the intestinal tract.
Proceeding from the lumen outwards to the serosa, the four layers of the intestinal tract visible during ultrasound examination are:
■ Mucosa (slightly hypoechoic)
■ Submucosa (hyperechoic)
■ Muscularis (hypoechoic)
■ Serosa (brightly hyperechoic)
In addition to the four physical layers described above, a very narrow hyperechoic layer is usually visible at the interface between the mucosa itself and the intestinal lumen.
5.1.2.1 Anatomical features of the small intestine
5.1.2.1.1 Increasing available surface area
There are several anatomical features that increase the available surface area of the intestinal epithelium (Figure 5.1).
The inward folding of the mucosa into grossly visible folds or rugae increases the available surface area by a factor of approximately 3 over a plain tube of equivalent diameter. The mucosa is arranged into finger-like structures called villi, which increase the surface area by an additional factor of approximately 10.
Enterocytes have a microscopic brush-border modification of their luminal cell membrane called microvilli. These microvilli increase the available surface area for digestion and absorption of nutrients by a factor of nearly 20 over the surface area of the villous projections.
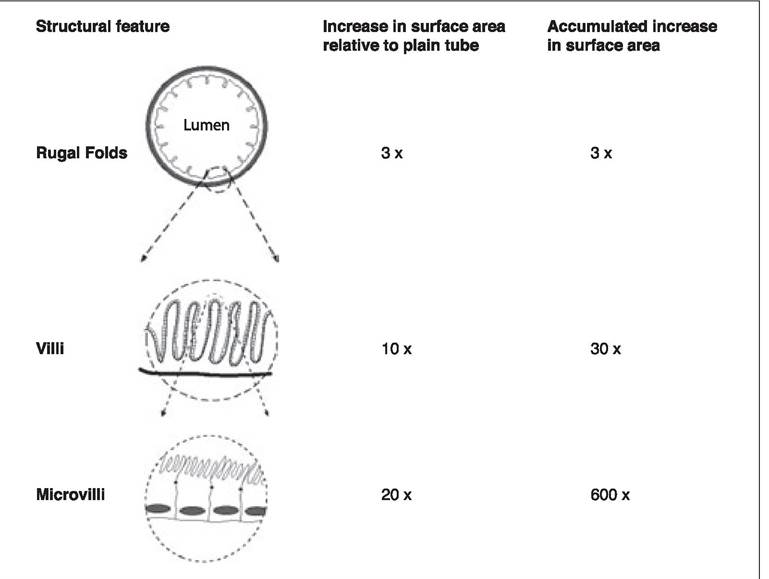
Figure 5.1:
Macroscopic and microscopic anatomical structures increasing the surface area of the intestine.
In combination the three structural elements of rugae, villi, and microvilli increase the effective surface area of the intestinal mucosa by a factor of approximately 600 compared to a plain tube of the same diameter. The loss of one element, for instance villous blunting, as occurs with some viral enteropathies or IBD, will dramatically alter the function of the small intestine.
5.1.2.1.2 Microscopic anatomy of the intestinal tract
The villus is the functional unit of the small intestinal mucosa. Enterocytes arise from the division of stem cells in crypts at the base of the villus. They then migrate and mature along the length of the villus and are finally shed from the villous tip into the lumen. As enterocytes migrate along the villus their function alters, starting from a mainly secretory function in the crypt and lower villus, maturing to a mostly absorptive function in the upper part of the villus (see 5.2.2).
5.1.2.1.3 Spatial variation in intestinal structure
The small intestine has three major anatomical divisions. Proceeding from oral to aboral, these are the duodenum, the jejunum, and the ileum. The specific function of these three regions varies, with digestion of nutrients primarily occurring in the duodenum. The jejunum has both transport and absorptive functions, and the ileum is almost exclusively absorptive. The size and structure of the villi vary down the length of the small intestine, with duodenal villi being short and blunt, while the villi of the jejunum are long and narrow. In the duodenum, the ratio of the villus length to crypt depth is nearly equal, while the jejunal and ileal villi are much longer than the crypt depth. This anatomical difference reflects the greater need for increased surface area for the absorptive processes in the jejunum and ileum.
gastric antral mucosal extracts on gastric acid secretion. The responsible substance was originally called “gastric secretin”, but was subsequently abbreviated to gastrin.3 Gastrin is a 17-amino- acid peptide, produced by “G” cells in the antral mucosa in response to the presence of protein and fat in the distal gastric antrum. Gastrin acts upon enterochromaffin-like cells to increase the production of histamine by these cells. Histamine released by the enterochromaffin-like cells then stimulates gastric acid production by the chief cells. Entry of food into the small intestine stimulates the production of cholecystokinin (CCK). CCK is released from duodenal “I” cells, stimulating the emptying of the gall bladder and the production of digestive enzymes by the pancreatic acinar cells.
These three “classical” GI hormones were isolated and their predominant functions identified in the early decades of the twentieth century. As all three are stimulatory, elucidation of their effects on the GI tract and associated organs was rela-
Table 5.1: Examples of endocrine compounds with major effects on the gastrointestinal tract
| Hormone | Source | Function(s) |
| "Classical" GI Hormones | ||
| Cholecystokinin | Duodenal mucosa, “I” cells | Stimulates gall-bladder contraction and pancreatic enzyme secretion |
| Gastrin | Gastric antrum, “G” cells | Stimulates gastric acid secretion |
| Secretin | Duodenal mucosa, “S” cells | Stimulates pancreatic bicarbonate secretion |
| "Modern" GI Hormones | ||
| Enteroglucagon | Distal small intestine, “L” Cells | Enhances release of insulin in response to glucose |
| Gastric Inhibitory Peptide | Proximal small intestine | Inhibits gastric motility and acid secretion |
| Ghrelin | Gastric epithelium | Stimulation of growth hormone release, regulation of appetite |
| Motilin | Proximal small intestine | Stimulates migrating motor complexes |
| Pancreatic Polypeptide | Pancreatic acinar cells | Affects pancreatic secretion, complex effects on GI motility |
| Somatostatin | Pancreas, enteric nervous system, GI epithelium | Inhibits secretion of many other GI hormones |
tively simple.
Inhibitory hormones, whose effects are manifest as decreases in GI secretion or motility, are more difficult to demonstrate in whole-animal studies, and the major hormone with inhibitory effects on the GI tract, somatostatin, was not isolated and identified until nearly forty years later.The small intestinal mucosa is the source of motilin, a peptide hormone that stimulates GI motility. Motilin is released into the circulation during the interdigestive phase. This pulsatile release of motilin stimulates cyclical muscle contractions that sweep the length of the intestine. These cyclical “housekeeping” contractions are called the migrating motor complex.
Sensations of hunger and satiety are important in the regulation of food intake. Ghrelin, another peptide hormone produced in the gastric mucosa, is released into the circulation when the stomach is empty. Ghrelin acts on the CNS to regulate hunger and satiety, with circulating concentrations increasing during the interdigestive period, which then rapidly decrease after feeding. Ghrelins name is derived from another important systemic function of this hormone, acting as a growth-hormone- releasing hormone (GhRH) in the pituitary. Ghrelin acts in concert with somatostatin and GhRH to regulate the release of growth hormone. Ghrelin is a potent stimulator of growth hormone release, while somatostatin is a suppressor of growth hormone release. The actions and release patterns of ghrelin and somatostatin, in concert with GhRH from the hypothalamus, lead to a highly pulsatile release of growth hormone. Thus, the stomach mucosa, and the state of GI filling, play an important role in the integrated regulation of body-wide physiology by its production and release of ghrelin, and hence its influence on growth hormone production.
Ghrelin concentrations are lower in obese humans than in lean controls. Conversely, in patients with Prader-Willi syndrome, extremely high circulating concentrations of ghrelin are observed in association with a severely increased, uncontrollable, voracious appetite and extreme obesity.4 Studies of ghrelin in companion animals are currently lacking, thus it is not known if similar changes occur in dogs or cats with obesity.
5.2.4