One species three names: a question of synonymy
I. ventalloi was first described by Gil Collado based on the morphological characters of a female tick parasitizing an Athene noctua captured in Barcellona [15]. The identification of this small-sized Ixodes, among other features, was based on the presence of particularly large and curved auriculae (Figure 1), differentiating it from the other European species, as the author wrote “Las auriculas de la base del capitulo, muy tipicas y de forma de “asta de touro” (...) la distinguen netamente de las especies europeas (...)” [15].
In the following years of its description, I. ventalloi was misclassified and as a consequence, confusion arose regarding the morphological features and the ecology of this tick. I. ventalloi was either incorrectly ascribed as a new species, I. thompsoni, or confounded with I. festai and used for the redescription of the latter species mistaking both entities, as detailed by Gilot and Perez [16]. I. festai, originally described by Rondelli in 1926 based on the analysis of a female specimen found parasitizing a Libyan Alectoris barbara, is a bird-associated tick contrasting with the more permissive nature of I. ventalloi, as revised bellow in this chapter. Several works mention I. thompsoni and I. festai sensu Arthur as a synonym of I. ventalloi, but the definitive validation of this species was only achieved with the studies of Gilot, Morel and Perez [16-18].Detailed descriptions of relevant morphological features have been subsequently updated, in some cases supported by illustrations and microscope images to better assist acarologists in I. ventalloi identification [14, 19-21]. Moreover, the application of mitochondrial DNA analysis using molecular targets, such as 12S rRNA, 16S rRNA, cytochrome c oxidase subunit 1 (cox1), has proven useful to complement the traditional morphological identification [12-14].
It also enabled the study of the population genetic structure. The molecular characterization of 92 I. ventalloi adults collected in cats from Lipari Island (Southern Italy) revealed the presence of a great genetic variability with the identification of eight haplotypes for 16S rRNA and 16 haplotypes for cox1, clustering in two sister clades—genogroup A, comprising 71% of the samples and genogroup B [13]. Interestingly, 16S rRNA sequences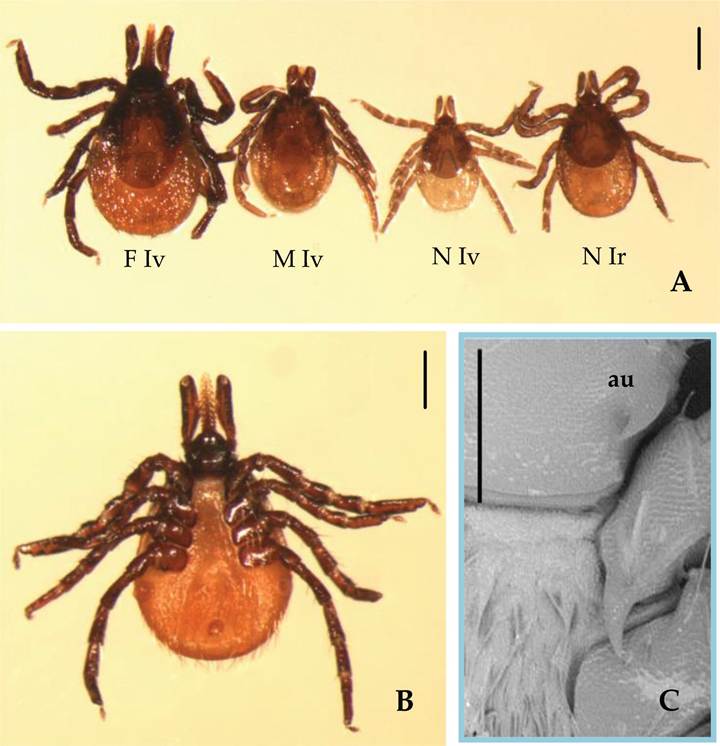
Figure 1.
Morphologicalfeatures of Ixodes ventalloi: (A) Size comparison of questing I. ventalloi female (F Iv), male (M Iv), and nymph (N Iv) with a questing Ixodes ricinus nymph (N Ir), dorsal view, scale bar 2 mm; (B) ventral view of a questing I. ventalloi female, scale bar 2 mm; (C) scanning electron microscopy detail of the recurved auriculae (au), the hallmark feature first pointed by Gill Collado [15]for differentiation from the other European Ixodes, scale bar 200 μm.
closest to those belonging to genogroup A were also documented in Spain and Portugal [12, 14]. In the latter study, 12S and 16S rRNA genes have been targeted in 48 questing I. ventalloi (nymphs and adults), resulting in the identification of six haplotypes (IvH1-H6) but with a low degree of nucleotide variation placing them all in Latrofas genogroup A [12]. Figure 2 represents the phylogenetic distance, based on 16S rRNA sequences, of the I. ventalloi specimens collected in Italy, Spain, and Portugal, comparing to those related (>91% homology) Ixodes species. These results highlight the need for further genetic characterization of I. ventalloi population, increasing both the molecular coverage and the number of studied specimens from other geographical origins.
The I. ventalloi sequences obtained in the aforementioned studies were deposited in GenBank under the accession numbers: KU178964-KU178979 for cox1; KU178956-KU178963, KY231931, MF370642-43, MF621228, MF621233, MG210719-20 for 16S rDNA; MF370631-32, MF621221, MF621226, MG210717-18 for 12S rDNA.
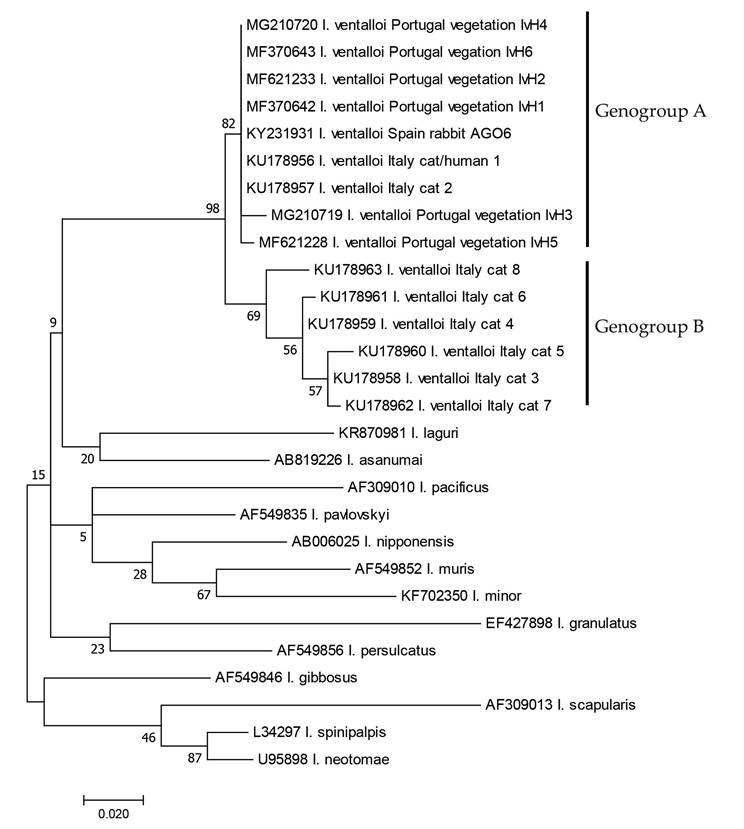
Figure 2.
Phylogenetic trees based on 16S rRNA sequences obtained from Ixodes ventalloi collected in Italy, Spain and Portugal, comparing to sequences of other related (>91% homology) Ixodes species available in GenBank. Phylogenetic relationships were assessed computing the maximum likelihood method on MEGA7 [22]. Best fitting substitution models were determined using MEGA7 model selection method. Phylogenetic tree was constructed using General time reversible model, modulated by using a discrete Gamma distribution (+G) and based on the analysis of 1000 replicates. All positions with less than 75% site coverage were eliminated. That is, fewer than 25% alignment gaps, missing data, and ambiguous bases were allowed at any position. There were a total of 247 positions in the final dataset. Accession numbers are followed by species name, and in some case the origin of sequences and haplotypes designation. Branch lengths represent the number of substitutions per site inferred according to the scale-bar.
3.