The Dynamics of the Epizootic
The global distribution of E. cancerae (de Hoog et al. 2011) suggests that it may have been present in Brazil prior to the beginning of the epizootic in the late 1990s. LCD outbreaks, with local crab mortality rates of up to 90%, may have taken place before.
Indeed elder fishermen from regions along the Brazilian coast more than 1500 km apart indicated that extensive mortalities of mangrove-land crabs took place more than 50 years ago. Assuming that the species has a global distribution, changes in host or in environmental conditions, rather than emergence of virulent fungal genotypes, are likely causes of the epizootic.The original habitat of the etiologic species of LCD, despite many attempts, remains unknown. Search efforts to identify habitats and eventual other hosts of E. cancerae and F. brasiliensis endemic areas of crab disease in the recent past were partially successful. Guerra et al. (2013), using techniques of fungus isolation and molecular detection on mangrove plant substrates and mud associated with crab burrows yielded positive results with F. brasiliensis in samples from plant branches and roots. However, E. cancerae was again not recovered from any environmental sample. Perhaps this species has a hitherto uncovered reservoir. During periods of the absence of LCD, E. cancerae was detected in specimens of U. cordatus and, in a single specimen of Cardisoma guanhumi (“guaiamum”), another Ocypodidae crab species (Boeger and Pie 2006). Although animals were positive for the agent of LCD, they were free of clinical signs of the disease. It was therefore concluded that E. cancerae was able to reside asymptomatically in the mangrove-land crab and eventually in its close relative C. guanhumi. Perhaps this specific lineage of E. cancerae is a true host-specific pathogen, requiring as yet unknown conditions to become fulminant.
In contrast, F. brasiliensis represents an opportunist that is capable of colonizing weakened, diseased crabs.Starting in 1997 from the first reported locality with high mortality in the mangrove land of Goiana in Pernambuco, putative LCD spread to northern and southern mangroves in a wave-like fashion (Fig. 11.2). Greater mortalities were usually seen during the first events (first wave) in each direction, with lower mortalities occurring in populations where the disease had occurred previously. Unsuitable environmental conditions are known to cause stress and subsequent weakening of the immune response of the host and making the crabs susceptible to infection (Stentiford and Feist 2005). Several studies reported that that relative immune competence may vary among individual mangrove-land crabs. Some individuals were apparently capable of impeding (at least temporarily) the invasion and multiplication of the fungus within their tissues. In some crabs that looked healthy, hyphae and yeast-like cells were found within hemocytic encapsulations and/or agglutinations only (Boeger and Pie 2006). In other crabs, however, rapid multiplication of fungal cells appeared to overwhelm their immune response, resulting in tissue invasion and dissemination leading to LCD. A small percentage of animals managed to survive events of LCD of mangrove-land crabs (Nobrega and Nishida 2003; Schmidt 2006). Part of these survivors may not have been exposed to the causative agents, but others may have been capable of clearing fungal invasion of their tissues. Genetic, physiological, and/or environmental factors may determine differences in strength of the immunological response to E. cancerae within a population of the mangrove-land crab; the relative contribution of these factors warrants further exploration.
The wave pattern observed in the mortality associated with LCD suggests that there may be an interplay between demographic and epidemiologic processes within each population. Schmidt (2006) suggested that the most likely dispersion pathway for the fungus of LCD is the marine environment, since he found crabs to present higher mortality rates in the lower intertidal zones.
Environmental processes interacted with each other; general pattern of mortalities and dispersion were reproduced in computational models by Ferreira et al. (2009) and Avila et al. (2012). However, Boeger and Pie (2006) suggested that the wave pattern of dispersion could be associated with a variation of the resistance of crabs along the year. As mortalities often occurred following the “andada” (period when crab leave their burrows to copulate), these authors suggested that the resistance of the crabs would be reduced following this period of apparent stress. The dispersion would promote extensive mortalities of mangrove-land crab population in the limits of distribution during the first year, but subsequent outbreaks would be less extensive due to the larger percentage of resistant individuals in the populations—since susceptible hosts had largely disappeared from the population during previous outbreaks.Curiously, the outbreaks spread north and southward, but in both directions reached a limit in distribution. The southernmost limit of dispersion of mortalities associated with LCD occurred in the mangroves of the state of Espirito Santo in the year 2006. Despite recurrent mortalities during the next 2 years in the region, LCD did not disperse toward mangrove populations located south of this state. Similarly, mortalities were reported in populations of mangrove-land crab in the northern states of Rio Grande do Norte and Ceara in 2000. The northernmost report occurred in the state of Piaui, in 2003. No mortalities were reported in the extensive mangroves located in the northern states of Maranhao and Para—the latter is the state that presents that largest concentration of mangroves in Brazil.
An in vitro experiment (Orelis-Ribeiro et al. 2017) showed that the agent of LCD, the black yeast E. cancerae was remarkably tolerant to variable salinity levels, supporting an extensive period of exposure (1 week) to the osmotic stress of 38 ppt salinity, which in nature is recorded only during dry periods of the Brazilian northeastern coast (Memery et al.
2000, F field 2005). Thus, the experimental results support the hypothesis of dispersion of the etiological agent through the marine environment in most Brazilian coastal areas. However, higher temperature (30 °C) caused a reduction of about 1.2-fold in colony-forming units. The maximum growth temperature of E. cancerae is 33 °C and optimal growth at 24-27 °C (de Hoog et al. 2011) (Fig. 11.6). Hence, it seems plausible to suggest that sea surface temperatures either above or below this optimum growth range apparently represent a key factor influencing the dispersion of LCD-related mortalities to the north-south limits of distribution of U. cordatus.Still, the factors associated particularly with the southern limits of dispersion of E. cancerae are not clear. It appears that the rarefaction in mangrove distribution in the southern coast of Espirito Santo and the northern coast of the state of Rio de Janeiro could represent a buffer area precluding the stepping-stone dispersion of the etiologic agent of LCD. The change in direction of the coast from a roughly northsouth to a northeast-southwestern, observed immediately southern of the limit of distribution of the outbreaks, increases the distance between the main coastal current from the coast which may further reduce the possibility for the dispersing fungal forms to reach the coastal mangroves.
Mathematical modeling has proven to be a valuable tool to providing insights on the spatiotemporal dynamics of LCD (Ferreira et al. 2009, Avila et al. 2012). In a first attempt to elaborate a model that reflects the periodicity and the observed fluctuations in the incidence of the mortality events within a mangrove area, Ferreira et al. (2009) separated the adult mangrove-land crab population into infected and susceptible groups. Noteworthy is that simulations over the range of parameter values successfully reproduced the cyclic pattern of the epizootic events observed in nature. Essentially, the model predicts that in areas where the mangrove-land crab population is more susceptible, or E.
cancerae is highly virulent, the incidence of high-profile mortality events is higher, and the cycles occur because the susceptible crabs are driven to low densities at which the black yeast population can no longer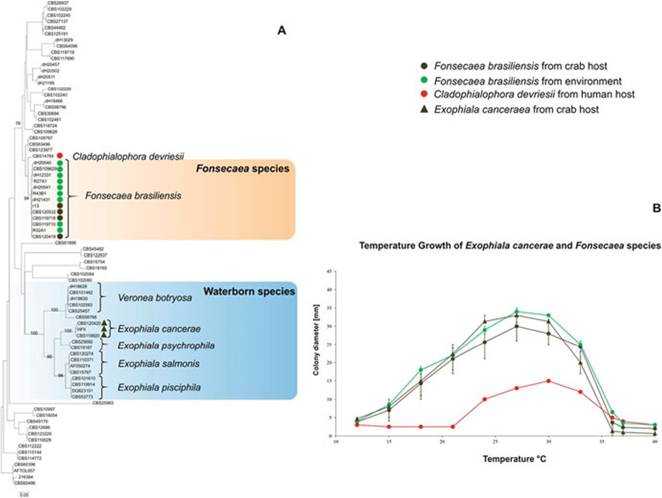
Fig. 11.6 Molecular ecology of the lethargic crab disease agents: (A) phylogenetic tree based on confidently aligned ITS sequences constructed with maximum likelihood implemented in RAxML. Colored rectangles indicate main clades recognized within by Vicente et al. (2012) and de Hoog et al. (2011) which grouped the two disease agents Fonsecaea brasiliensis and Exophiala cancaerae, respectively; (B) Temperature relations of F. brasiliensis, E. cancerae and Cladophialophora devriesii. For interpretation environmental strains (in green), strains isolated from crabs (in brown), clinical strain from human patient (in red)
spread, therefore also decreasing. When the susceptible crab population recovers to a threshold, another epidemic peak emerges, and so on. In summary, although stress associated with mating season has been previously suggested as a possible factor influencing the dynamic of the outbreaks, the developed model shows that the oscillations may be a consequence of the relationship between demographic and epidemiological parameters. Furthermore, considering the implications of this model, the authors hypothesized that the introduction of resistant mangrove-land crabs or the sustainable management of its population through capture could represent effective steps toward preventing the spread of LCD. Adding another layer of complexity to the previous model, Avila et al. (2012) formulated a model to study the propagation of LCD between mangroves as a result of dispersion of E. cancerae in sea (either by diffusion or ocean currents). The study assessed different conditions for the existence of epidemic waves, thus determining the minimum speed at which they travel. This model pointed out the existence of traveling wave solutions, which strengthened the previous hypotheses regarding strategies for disease control. Simulations on the study of the speed of the epidemic wave indicated that ceasing the collection of mangrove-land crabs in estuaries close to the LCD-affected areas will increase disease dispersion throughout the Brazilian coast. In addition, the results also suggested that spread of the epizootic events mainly from the north to south could be owing to the influence of ocean currents in the dispersion of the fungus.
The reports of mortalities of populations of the mangrove-land crab are presently scarce and limited to mangroves located in the southernmost limit known for the outbreaks, in the state of Espirito Santo and southern Bahia.