A vector’s potential neglected or negligible?
During several years, information regarding I. ventalloi pathobiome was almost absent. The first association of this tick to a potential tick-borne agent was reported in 1984 by Chastel et al.
[47]. In this study, strains of the coltivirus Eyach (EYAV), a virus belonging to Colorado tick fever group, were isolated from both I. ventalloi and I. ricinus ticks that were found parasitizing a wild rabbit in Northwestern France. Eyach virus was previously described in I. ricinus from West Germany, subsequently found on several wild mammals and indirectly linked to patients with neurological disorders, as tick-borne encephalitis, polyradiculoneuritis, and meningopolyneuritis, on a base of serology [48]. In 2004, we have also reported I. ventalloi infection by Anaplasma phagocytophilum, a species with variant strains implicated in human and domestic animal cases of granulocytic anaplasmosis [8]. The growing interest on I. ventalloi observed in the last 10 years has resulted in an increasing number of papers and the detection of diverse microorganisms associated to this tick species. Table 2 resumes the microbial agents that have been found in I. ventalloi, providing information on ticks stage, sex, and molecular identification (haplotypes), when available. Overall, 13 agents have already been associated to I. ventalloi, and infected ticks were found feeding on wild animals, as well as on domestic cats and on a human, pointing for a potential role as vector that might have both medical and veterinary implications.Another justification for the presence of agent’s nucleic acids in parasitizing ticks can also be the presence of host-infected blood in arthropods' midgut rather than a true vector potential. However, it is worthy of note that some of the I. ventalloi positives were indeed unfed ticks, as detailed. The first record dates back to 2004 when the authors were investigating A.
phagocytophilum in I. ricinus and their sympatric ticks in Setubal District, Portugal [8]. The screened sites were mainly suburban wooded areas in some cases used for grazing and with evidence| Microorganism | Ticks origin | Ticks haplotypes§ | Reference |
| Virus | |||
| Coltivirus Eyach | Oryctolagus cuniculus | ND | [47] |
| Bacteria | |||
| Anaplasma marginale | Vegetation | ND | [49] |
| A. phagocytophilum | Vegetation, Felis catus | ND | [8, 10] |
| Vegetation | IvH1, IvH3, IvH5 | [12] | |
| Ehrlichia canis | Felis catus | ND | [36] |
| Ca Neoehrlichia sp. | Vegetation | ND | [12] |
| Rickettsia helvetica | Asio flammeus | ND | [9] |
| Lynx pardinus, Vulpes vulpes | ND | [38] | |
| Felis catus, Rallus aquaticus | ND | [32] | |
| Felis catus* | ND | [36] | |
| Homo sapiens | Haplotype 1 | [13, 37] | |
| R. monacensis | Oryctolagus cuniculus | ND | [29] |
| Lynx pardinus, Vulpes vulpes, Genetta genetta | ND | [38] | |
| Felis catus* | ND | [32, 36] | |
| Coxiella burnetii | Alectoris chukar, Lepus europaeus | ND | [31] |
| Vegetation | IvH2, IvH3, IvH5, IvH6 | [12] | |
| Bartonella clarridgeiae | Felis catus⅛ | ND | [36] |
| Borrelia valaisiana | Felis catus, Rallus aquaticus, Phasianus colchicus* | ND | [32] |
| B. spielmanii | Felis catus, Phasianus colchicusY | ND | [32] |
| Protozoa | |||
| Leishmania infantum | Felis catus⅛ | ND | [36] |
| Theileria annulata | Vegetation | ND | [49] |
*One tick co-infected with Rickettsia Helvetica and R. monacensis [36].
^Onefemale tick coinfected with Bartonella clarridgeiae and Leishmania infantum.
¥One tick co-infected with Borrelia valaisiana and B. spielmanii.
sAll these haplotypes have been identified as belonging togenogroup A and were submitted to Genbank under the accession numbers: Haplotype 1, KU178956 (16S rDNA) and KU178964 (cox1); IvH1, MF370631 (12S rDNA) and MF370642 (16S rDNA); IvH2, MF621226 (12S rDNA) and MF621233 (16S rDNA); IvH3, MG210717 (12S rDNA) and MG210719 (16S rDNA); IvH5, MF621221 (12S rDNA) and MF621228 (16S rDNA); IvH6, MF370632 (12S rDNA) and MF370643 (16S rDNA), as previously described [12, 13].
Table 2.
List of microorganisms and parasitesfound in Ixodeis ventalloi.
of wild animals presence, as rabbits. Out of the 93 I. ventalloi collected, A. phago- cytophilum was recorded in a questing nymph and also in a male found attached to a free-roaming cat. The sequences obtained from both ticks were found to be a new A. phagocytophilum variant based ongroEL and msp2 gene analysis [8]. This finding was subsequently reinforced by the detection of the same A. phagocyto- philum variant also in an I. ventalloi female feeding on a cat from another district of mainland, Santarem District [10]. More recently, a retrospective study using the DNA material stored from Setubal district ticks also resulted in the detection of questing I. ventalloi specimens of Anaplasma marginale (four nymphs and one male) and Theileria annulata (one female) [49]. New data that link more agents to questing I. ventalloi were submitted for publication in the beginning of 2018 [12]. That study was undertaken in Parque Florestal de Monsanto (PFM), a recreational area located in the urban perimeter of Lisbon city. This is a highly used park for petting and several outdoor activities. Overall, eight tick species were found questing in PFM with a preponderance of I. ventalloi. A preliminary 1-year screening to define the best season for collection, established that the period of activity for this species extended from November to June, with a peak in spring (Figure 3) [50]. Interesting, both of these findings (abundance and seasonality) were reinforced in Torina et al. comprehensive study [45]. In our case, the diversity of PFM ticks and the particular abundance of I. ventalloi were attributed to the park's wild population, composed of over 100 species of small mammals and birds [51].
Regarding tick-borne agents, questing I. ventalloi in PFM were found harboring A. phagocytophilum (two males, one female, and two nymphs), Coxiella burnetii (five males, three females, and one nymph), and a potentially new agent
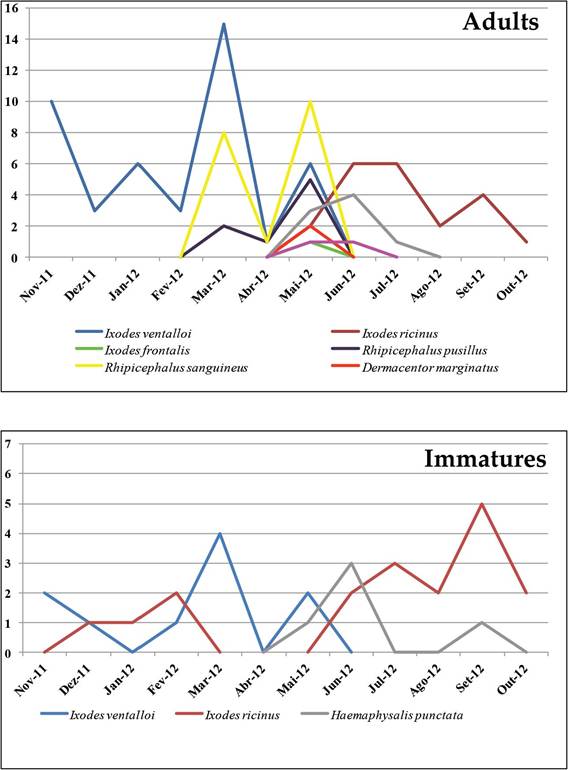
Figure 3.
The distribution of immature and adult tick species collected in PFM during 1-year period (2011-2012) [50].
close related to Candidatus (Ca.) Neoehrlichia mikurensis (one female and one male). Interestingly, two A. phagocytophilum variants were detected [12]. The more representative was a new variant of A. phagocytophilum previously detected in both Setubal and Santarem districts (and here found on four ticks) [8, 10]. This reinforces previous data sustaining a divergent variant of A. phagocytophilum, not clustering in none of the four ecotypes defined by Jahfari et al. [52], with the closest sequence sharing only 95% homology and belonging to ecotype IV that is composed of sequences of the agent derived from birds. Another A. phagocytophilum variant was obtained from a single I. ventalloi, clustering the ecotype I that is composed by agent’s sequences associated to human and domestic animal cases of granulocytic anaplasmosis [52]. It was also worth of mention that positive ticks were found questing in different occasions showing the existence of active cycles for these agents in PFM [12]. The molecular identification of nine positive ticks confirmed that all belonged to Latrofa’s genotype A, based on 16S rDNA analysis. The obtained haplotypes and the GenBank accession numbers are presented in Table 2.
In all the aforementioned Portuguese areas, infected I. ventalloi were found questing along with other moisture-demanding ticks, as the mega-vector I. ricinus. Both tick species are considered sympatric sharing geographical distribution, vertebrate hosts, and possible their agents [11]. The presence of alternate ticks (generally endophilic ticks) has been associated to the existence of secondary maintenance cycles for some Ixodes -borne agents [53]. If I. ventalloi has such a role and thus contributes indirectly to the occurrence of I. ricinus-borne diseases is yet to be investigated.
5.