ADRENAL GLANDS
1. Where are the adrenal glands located?
2. What are the two principal hormones of the adrenal cortex? What is their biochemical classification?
3. What is the role of the glucocorticoids in carbohydrate metabolism? What is the main noncarbohydrate source of new glucose formation?
4.
Do glucocorticoids have some mineralocorticoid activity?5. What is the principal function of the mineralocorticoids? Do they possess some glucocorticoid activity?
6. What regulates the secretion of the glucocorticoids?
7. What are the processes by which aldosterone secretion increases?
8. What are the hormones of the adrenal medulla?
9. What is the biochemical classification for epinephrine and norepinephrine? Are they also considered catecholamines?
0. What division of the autonomic nervous system secretes norepinephrine? Is it a postganglionic or preganglionic secretion?
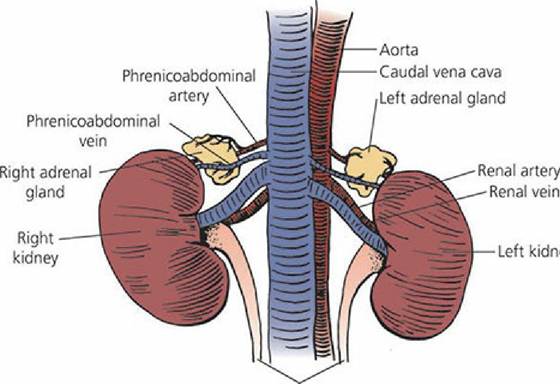
The adrenal glands are small, paired structures that lie immediately cranial to the kidneys and are close to the junction of the renal vein with the caudal vena cava (Figure 6-5). A sagittal section of the adrenal gland (Figure 6-6) shows an outer cortex and an inner medulla. The adrenal cortex has three distinct cell types: arranged.in zones from the outside to the inside,.the zona glomerulosa, zona fasciculata, and zona reticularis. The adrenal medulla is homogeneous in structure and contains secretory granules. Its nerve supply is by way of preganglionic sympathetic neurons. The cells of the medulla are thought to be modified postganglionic sympathetic nerve cell bodies.
Ureters
■ FIGURE 6-5 The canine adrenal glands (ventral view). Their blood supply and venous drainage is by way of branches from the phrenicoabdominal arteries and veins.
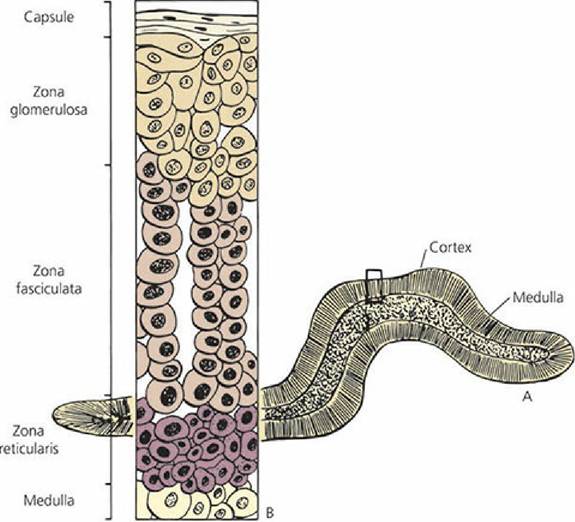
■ FIGURE 6-6 Diagrammatic representation of the adrenal gland.
A. Cross-section of the adrenal gland showing the contrasting appearance of the cortex and medulla. B. Magnification of boxed-in area in A that shows the different cell types associated with the three zones of the cortex.Hormones of the Adrenal Cortex
The hormones of the adrenal cortex are steroids formed mainly from cholesterol. The membrane of the adrenal cortex has receptors for low-density lipoproteins (rich in cholesterol) and, after their attachment, these are absorbed by endocytosis. Seven adrenocortical hormones (corticosteroids) are recognized as secretions of the adrenal cortex. Four of these - corticosterone, cortisol, cortisone, and 11-dehydrocorticosterone -.are termed glucocorticoids. The other three - 11- deoxycorticosterone, 17-hydroxy-,11-deoxycorticosterone, and aldosterone - are called mineralocorticoids. The structural formulas of the two principal adrenocortical steroids (aldosterone and cortisol) are presented in Figure 6-7.
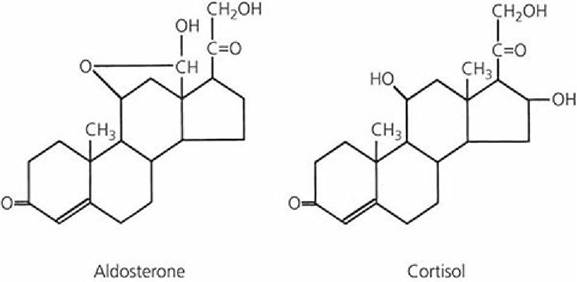
■ FIGURE 6-7 Structural formulas of the principal adrenocortical hormones.
Glucocorticoid Functions and Regulations
The glucocorticoids have a principal role in carbohydrate metabolism in that they enhance gluconeogenesis. The noncarbohydrate source from which new glucose is synthesized is mostly protein, but a definite effect on fat metabolism is also recognized. Two other hormones, glucagon and epinephrine, increase blood glucose levels by glycolysis of liver glycogen. The glucocorticoids, however, seem to be necessary for glycolysis affected by glucagon and epinephrine. The gluconeogenic effect of the glucocorticoids is the basis for their use in the treatment of bovine ketosis (see Chapter 12).
A common therapeutic use of the glucocorticoids is related to their anti-inflammatory activity; they are included in ophthalmic preparations, otic (ear) drops, and skin ointments. Injection of glucocorticoids into inflamed articulations or bursae provides temporary relief and their systemic use is sometimes useful in the alleviation of some allergic responses.
Prolonged systemic use is often associated with exaggeration of other physiologic functions of the glucocorticoids, such as sodium retention related to their having some mineralocorticoid activity (retention of Na+ and H2O). More potent pharmacologic preparations (which require smaller doses) can minimize these side effects, but do not eliminate them altogether.The glucocorticoids are secreted by the zona fasciculata of the adrenal cortex. Their secretion is regulated by ACTH from the anterior pituitary. Free plasma cortisol (not protein-bound) concentrations influence ACTH secretion - low levels stimulate ACTH release and subsequent secretion of glucocorticoids from the zona fasciculata. Stimuli such as stress can also cause ACTH secretion that increases glucocorticoid concentrations above normal levels. An example of an adrenal response to adaptation can be seen with the overcrowding of domestic chickens whereby excess ACTH secretion results in adrenal hypertrophy because of greater output of glucocorticoids. The same phenomenon is observed in wild mammals whose population density increases.
Mineralocorticoid Functions and Regulation
The principal function of the mineralocorticoids can be illustrated by aldosterone and its action on the kidney to enhance sodium reabsorption and potassium excretion (see Chapter 11). The mineralocorticoids are also effective in promoting membrane transport in sweat glands, salivary glands, and intestinal mucosa, and between the intracellular and extracellular fluid compartments. Just as glucocorticoids have some mineralocorticoid activity, the mineralocorticoids have some glucocorticoid activity. These secondary activities are most apparent when therapeutic uses are made of these compounds and the amounts used are in excess of normal endogenous amounts.
Three processes are usually considered to be the means by which aldosterone secretion from the zona glomerulosa increases: (1) renin-angiotensin system; (2) increased plasma concentration of potassium (hyperkalemia); and (3) ACTH stimulation.
In the renin-angiotensin system (see Chapter 11), renin is secreted by juxtaglomerular cells in the kidney in response to their decreased perfusion by blood. Renin acts on a circulating blood globulin, angiotensinogen, to form angiotensinI. Angiotensin I is converted by the vascular endothelium to angiotensin II, which is the stimulus for the secretion of aldosterone from the zona glomerulosa. The result of this stimulation is promotion of Na+ reabsorption and consequent retention of water, which expands blood volume and thus reestablishes normal blood pressure (low blood pressure was the cause for renin secretion). Systemically, angiotensin II causes arteriolar vasoconstriction, whereby vascular resistance and systemic blood pressure are increased.
The secretion of aldosterone in response to hyperkalemia provides a means for controlling the critical plasma concentration of potassium (see Chapter 11). Aldosterone secretion promotes Na+ reabsorption with simultaneous K+ excretion. This action of aldosterone occurs in the distal tubule, collecting tubule, and collecting duct. At other places in the nephron, K+ is reabsorbed. Although Na+ is reabsorbed in the process of K+ excretion, the Na+ concentration in the plasma is not regulated by aldosterone. A decrease in plasma Na+ concentration can cause aldosterone secretion, but the decreases necessary for stimulation are of greater magnitude than the effective increases of K+ that cause aldosterone secretion.
The role of ACTH in promoting aldosterone secretion is of lesser significance. The increase in ACTH associated with stress causes some increased output of aldosterone and might augment the output produced by other means, such as angiotensin II.
Hormones of the Adrenal Medulla
The hormones of the adrenal medulla belong to the amine chemical class and are known as epinephrine (adrenaline) and norepinephrine (noradrenaline).
They are referred to as catecholamines and are derived from the amino acid tyrosine. The catecholamine hormones (including epinephrine and norepinephrine) are shown in Figure 6-8. Epinephrine is secreted only by the adrenal medulla, but norepinephrine is also secreted by postganglionic sympathetic neurons. More epinephrine is secreted by the adrenal medulla than norepinephrine. The inactivation of the catecholamines is rapid - the half-life of epinephrine is about 20 to 40 seconds.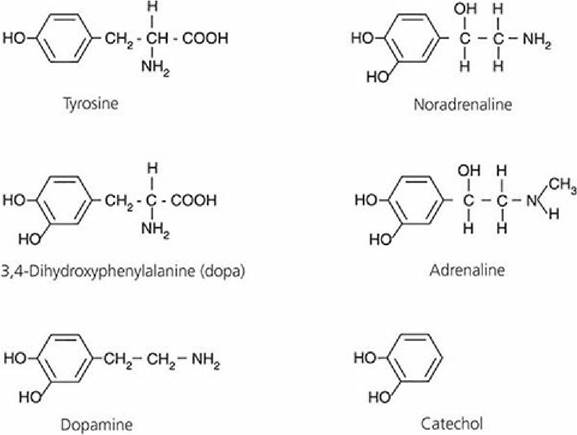
■ FIGURE 6-8 Structural formulas of catecholamine hormones. They are formed from the amino acid tyrosine and are derivatives of catechol. The abbreviation “dopa” is derived from the German name of this compound, dioxyphenylalanine.
It seems that the adrenal medullary secretion is a continuous process and increases dramatically during an emergency. The continuous secretion enables the maintenance of a state of readiness or tone, and the larger outpouring provides for an immediate response to emergencies.
The actions of epinephrine and norepinephrine are similar and differences are expressed depending on the receptors, which can have a preference for epinephrine or norepinephrine (see Chapter 4). The two adrenergic receptors are alpha and beta receptors. Alpha receptors are stimulatory (except in intestinal smooth muscle, where they are inhibitory) and beta receptors,are inhibitory (except those in cardiac muscle, where they are stimulatory). Epinephrine and norepinephrine stimulate both receptors, but the alpha effect of norepinephrine is more potent than that of epinephrine, and epinephrine has a more potent action than norepinephrine on the beta receptors.
In addition to the “fight-fright-flight” reactions associated with the catecholamines, they have pronounced metabolic effects. These are associated with the increased activity caused by catecholamines and include hyperglycemia, increased calorigenesis, lipolysis, and an elevated blood lactate concentration. The hyperglycemia results from enhanced liver glycogenolysis and the increased blood lactate level are caused by stimulation of muscle glycogenolysis. The calorigenic effect results from the increased muscle activity and an increase in lactic acid oxidation in the liver.
■