Development of the CNS
Formation of the neural tube
The notochord is derived from the mesoderm and establishes the cranial-caudal axis of the embryo. It induces the overlying ectoderm to become neurectoderm.
The neurectoderm thickens to form the neural plate along the dorsal, longitudinal axis of the embryo. A longitudinal trough appears in the midline of the neural plate, forming the neural groove. The sides of the plate rise up and seal over the top of the groove to form the neural tube, which surrounds a fluid-filled, neural canal. Closure of the neural tube is called primary neurulation; it occurs at about 20 days in the canine fetus (canine gestation is approximately 60 days). Closure starts in the cervical region and extends rostrally and caudally from that point. The ends of the tube may remain open for a period as the rostral and caudal neuropores. Just prior to neural tube formation, neural crest cells develop, located dorsolaterally at the junction between the neurectoderm and the ectoderm. The neural crest cells separate from the neurectoderm and the ectoderm as the tube develops. Columns of neural crest cells form along the dorsolateral aspects of the neural tube, while the ectoderm fuses dorsal to the neural tube creating the overlying skin. The neural tube extends the length of the embryo including into the head region and forms the basis of the spinal cord and the brain. The neural canal will develop into the inner ventricular system (see Chapter 3). Inside the head, the neural tube develops into three regions, the prosencephalon (forebrain = telencephalon and diencephalon), the mesencephalon (midbrain) and the rhombencephalon (hindbrain = metencephalon and myelencephalon). The sulcus limitans is a groove that forms in the lateral walls of the neural canal. It demarcates the tube into dorsal/alar and ventral/basal plates that are associated with sensory and motor functions, respectively (Fig. 2.1).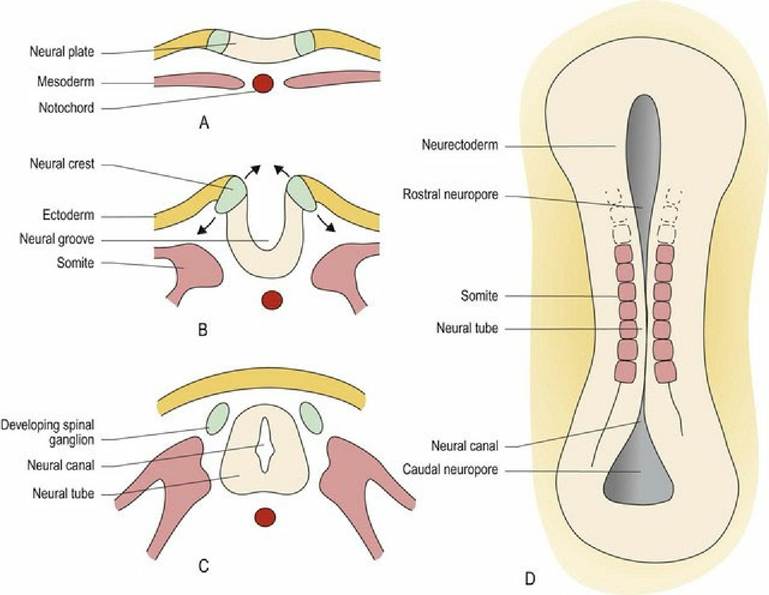
Fig. 2.1 The developing neural tube. (A-C) Transverse sections at sequential developmental stages. (D) Dorsal aspect, without overlying ectoderm, after neurulation.
During differentiation of the neural tube, three cell layers develop from the original pseudostratified, single cell layer. Innermost, sited around the neural canal, is the germinal layer, in which cellular proliferation occurs giving rise to both neurons and the supporting glial cells, oligodendrocytes and astrocytes. The postmitotic cells migrate so that the neurons are located in the middle or mantle layer, while their axons extend
into the marginal layer (Fig. 2.2). Astrocytes are found in both the mantle and marginal layer, while oligodendrocytes settle in the marginal layer to myelinate the axons. After development has finished the germinal layer becomes the ependymal layer surrounding the ventricular system of the brain and the spinal cord. The mantle layer becomes the grey matter and the marginal layer becomes the white matter. The grey and white matter form continuous columns; these extend the length of the spinal cord and into the brainstem. Overall, the brainstem develops from the neural tube in a similar way to the spinal cord (see Figs. 1.7, 10.2). However, the columns of grey and white matter become fragmented with the grey matter forming nuclei. Additionally, in the rhombencephalon, the dorsal aspect of the tube opens back up forming a groove-like structure, but with a thin cover called the medullary velum (velum - L = awning). The neural canal is expanded to become the fourth ventricle. The tube reforms in the midbrain, with a small-diameter neural canal, the mesencephalic aqueduct (Fig. 1.7).
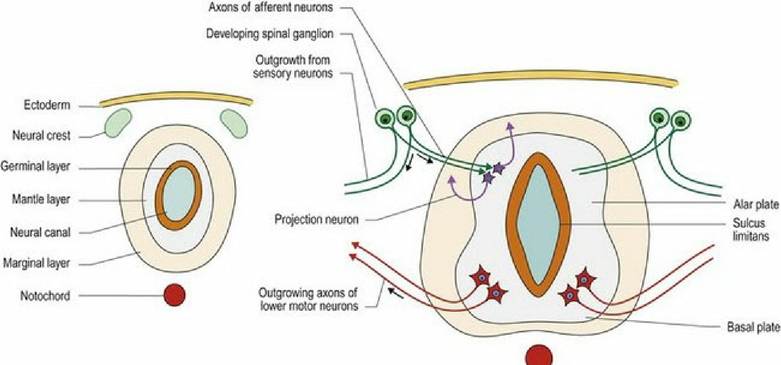
Fig. 2.2 Cellular differentiation of the neural tube. Arrows indicate direction of growth of the neuronal
processes.
Development of the brain
The cerebellum and the cerebral hemispheres develop as dorsal outgrowths of the neural tube.
The cerebellum forms from the dorsal aspect of the metencephalon arising as bilateral, rhombic lips that grow dorsally and fuse. Neurons arising from the germinal layer surrounding the future fourth ventricle migrate dorsally into the developing cerebellum to form the Purkinje cells and the neurons of the deep cerebellar nuclei. The external germinal layer is a second proliferative zone for the cerebellum. It develops on the dorsal, superficial aspect of the cerebellum and is the source of the remaining neurons for the cerebellum. The timing of cell proliferation in the external germinal layer is species specific. It may be complete before birth as in precocial animals such as many prey species. Or it continues postnatally, as in altricial animals, such as carnivores, and some prey species, such as rabbits and mice. An animal’s ability to ambulate in the perinatal period is directly related to the degree of cerebellar development. Prey species born in the open environment need to be able to get up and run with the herd immediately. Altricial species are usually born into a protective environment, such as a den or burrow and brain development can continue after birth (see Fig. 7.5).The forebrain develops as two regions, the diencephalon and the telencephalon (cerebrum). The thalamus of the diencephalon represents the rostral end of the neural tube and hence, the neural canal. Proliferation of tissue across the midline joins the walls of the neural tube and forms the interthalamic adhesion that splits the neural canal into dorsal and ventral components. These canals reunite rostral to the interthalamic adhesion, thus forming a vertically oriented, ring-like structure, the third ventricle. Bilateral outgrowths from the ventral thalamus form the optic cups and give rise to the neurons of the retina and the optic nerve. An outgrowth from the ventral midline joins a dorsal outgrowth from the future oral cavity, to form the hypophysis, comprising the neuro- and adenohypophysis, respectively.
The cerebral hemispheres arise from the dorsal aspect of the thalamus as massive, bilateral outgrowths. Each one is associated with a dorsolateral recess of the neural canal forming the lateral ventricles. These ventricles retain a connection to the third ventricle of the thalamus via the interventricular foramens. The germinal layer, which is still sited adjacent to the ventricles, is called the ventricular zone. It may continue to be a source of new neurons well into the postnatal period. Neurons migrate from the ventricular zone to the periphery of the developing cerebrum and their axons grow centrally from the surface. Thus much of the grey matter in the hemispheres is located superficially in the cerebral cortex with the white matter being located deep to that; this is the opposite of the spinal cord arrangement. However, some grey matter, such as the septal and basal nuclei and the hippocampus, develops deep within the cerebral hemispheres, adjacent to the lateral ventricles. In many species, the massive migration of neurons to the surface results in the cortical surface becoming convoluted and thrown into folds comprising gyri (ridges) and sulci (troughs). Some animals, such as rodents, lagomorphs and birds retain a smooth cerebral cortex; these species are called lissencephalic (see Fig. 1.8).
Molecular basis of differentiation
Differentiation of the neural tube is based on the expression of different genes and occurs in all three planes - ventrodorsal, mediolateral and craniocaudal (Fig. 2.3A-C). Ventrodorsally, two major morphogens affect differentiation of the neural tube. Sonic Hedgehog (SHH) protein arising from the notochord and subsequently the floor plate, causes nerve cells in the ventral aspect of the cord to differentiate into motor neurons. Bone Morphogenetic Protein (BMP) from the overlying ectoderm, and then the roof plate, causes cells in the dorsal aspect to differentiate into sensory neurons. Diffusion of these morphogens sets up dorsoventral and mediolateral concentration gradients.
Thus cells at different locations on the x-y grid will be exposed to different concentrations of morphogen, thereby determining their differentiation. In the craniocaudal direction the homeobox genes determine the regional specification along the neural tube that forms the spinal cord. There are many other genes involved in differentiation of the brain. It is also recognised now that astrocyte differentiation is regionally specified in a similar manner.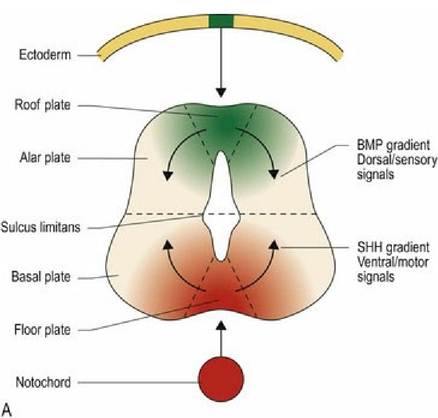
Fig. 2.3A Dorsoventral differentiation of the neural tube.
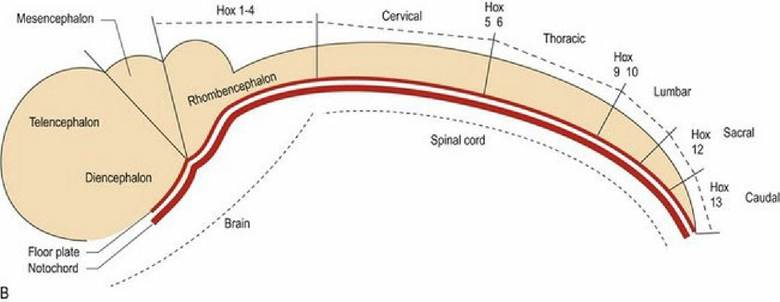
Fig. 2.3B Craniocaudal differentiation of the neural tube is determined by homeobox genes.
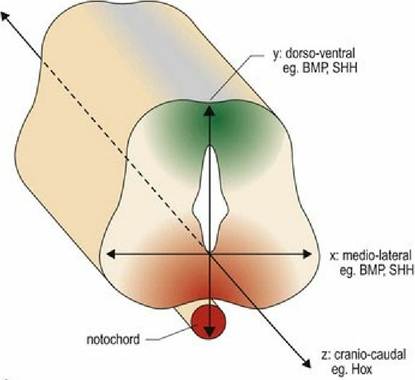
Fig. 2.3C Three-dimensional gradient of differentiation of the neural tube.
Glial cells lay down cellular scaffolds that assist migration of neuronal precursor cells to their final position in the developing cerebellum and the cerebrum. The process by which the neurons travel along the glial fibres involves recognition of specific proteins, such as astrotactin or reelin, which are expressed by glial or neuronal cells.
Neurons must connect to their appropriate target. The tips of axons arising from differentiating neurons navigate through the developing CNS by responding to molecular signals that may either attract or repel the tip (Fig. 2.4).
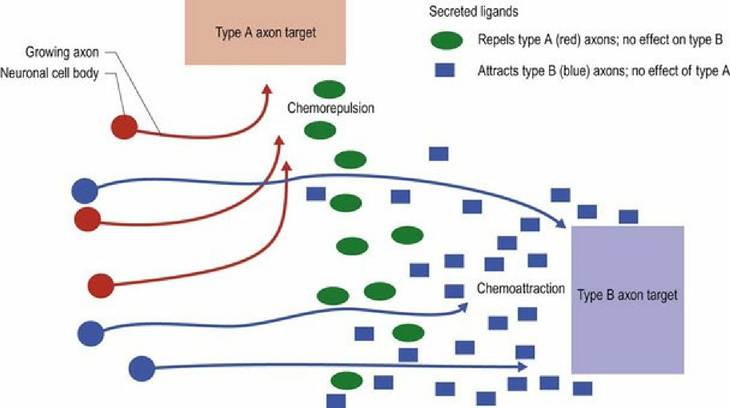
Fig. 2.4 Growth of axons in the developing nervous system is guided by molecular cues that attract or repel them.