INTRODUCTION
Many chemical messengers have been recognized for their role in facilitating communication between cells to achieve a specific homeostatic or homeorhetic response. The first system functions as a rapid channel, while the second one operates more gradually to produce a lasting reaction.
The endocrine system consists of various ductless endocrine glands that produce hormones, a specific type of chemical messengers.19.1.1 Glands and Classification
Glands are a collection of cells that are both structurally and functionally arranged to collaborate in producing a substance, such as an enzyme, sweat, saliva, milk, or hormone, and releasing it into either a duct or the bloodstream. Whether or not a duct is present is commonly used to categorize various glands into three main classes:
1. Endocrine glands: Secretions from ductless glands are frequently released directly into the blood. For example: T3/T4, Somatostatin, GH
2. Exocrine glands: Comprised of secretory acini that release their substances into a specific duct. For example: Bile, Saliva and etc.
3. Mixed glands: Glands that carry out both exocrine and endocrine roles. For example: Pancreas (Exo: enzymes, Endo: insulin), Testes (Exo: spermatozoa, Endo: testosterone)
19.1.2 Hormone Properties
Hormones are substances produced by specific endocrine glands or cells in small amounts, released into the bloodstream or interstitial fluid to reach target cells. This definition includes all hormonal chemical messengers, regardless of where they come from or how they work. Important traits of hormones include:
1. Specificity of receptors: Every hormone interacts with a particular high-affinity receptor, resulting in the formation of a hormone-receptor complex (HRC) that initiates a response in the target. The concentration of receptors differs between target cells and can be adjusted in response to hormone levels, known as upregulation or downregulation.
This active communication functions as enhancers and filters of hormone signaling.
2. Slow onset of action: Hormones generally take a long time to start showing biological effects, usually requiring hours to days, as they affect genetic processes to kickstart the transcription of effector genes.
3. Absence of enzyme function: Hormones do not have inherent enzyme function and are unable to directly accelerate intracellular enzymatic reactions.
4. Signal transduction: Hormones trigger intracellular cascades of reactions by acting as “first messengers” when they bind to target cell receptors. This frequently results in the formation of intracellular “second messengers.”
5. Metabolic clearance rate: Once hormones have produced effects in the body, they are either deactivated by the tissues they target or removed from the bloodstream through the excretory functions of the liver and kidneys. The elimination rate is opposite to their binding strength to carrier plasma proteins.
6. Feedback mechanisms: Feedback mechanisms involve close regulation of hormone secretion and concentration, mainly through negative feedback mechanisms, maintaining a narrow range.
19.1.3 Hormone Classification
Different hormones can be classified based on criteria such as where they come from, what chemicals they are made of, what they do in the body, how they work, and how easily they dissolve.
1. Their secretion source:
Classified as pituitary and non-pituitary hormones based on their site of production.
a. Pituitary hormones: Synthesized and secreted from different lobes of the pituitary gland. For example: GH, prolactin (PRL), LH, FSH, ACTH, MSH, ADH, and oxytocin.
b. Non-pituitary hormones: Include hormones produced by other endocrine glands except for the pituitary gland. For example: Somatocrinin (from the hypothalamus), insulin (from the pancreas), aldosterone (from the adrenal
DOI: 10.1201/9781003426851-19
179
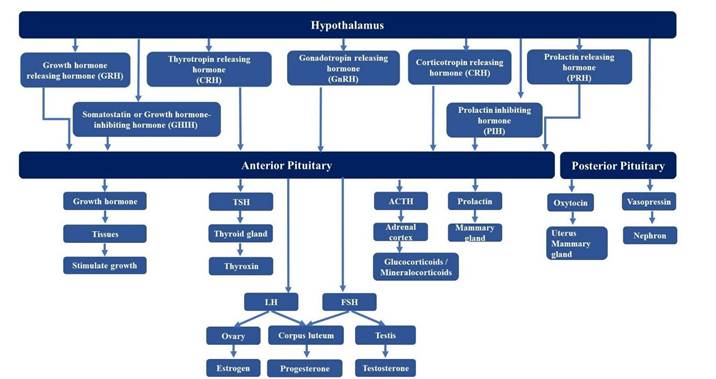
FIGURE 19.1 Classification of hormones in body.
cortex), T3/T4 (from the thyroid gland), etc (see Figure 19.1).
2. Chemical nature:
Different types of hormones can be classified based on their biochemical structure.
a. Protein hormones: These are composed of more than 50 amino acids. Protein hormones can be further categorized into:
i. Simple protein hormones (GH, Insulin, PRL, etc).
ii. Glycoprotein hormones (LH, FSH, TSH, and chorionic gonadotropins (eCG/hCG))
b. Polypeptide hormones: These consist of fewer than 50 amino acid residues. Examples include Calcitonin, TRH, Glucagon, etc.
c. Amine hormones: These are derived from amino acids through decarboxylation.
d. Steroid hormones: These are derived from cholesterol. They are primarily produced by gonads (e.g., estrogen, testosterone) and the adrenal cortex (e.g., Aldosterone, Cortisol).
e. Fatty acid derivatives: These hormones are synthesized from fatty acids. An example is the prostaglandin family (e.g., PGF2α, PGE2) derived from arachidonic acid (see Figure 19.1).
3. Their physiological action:
The integration of endocrine and nervous systems is crucial in controlling most basic physiological events such as growth, reproduction, intermediary metabolism, stress response (fight or flight response, emotional/physical stress, environmental stress), feeding responses, and water and electrolyte balance. Hence, the appropriate functioning of the endocrine system is vital for animal survival, optimum reproduction, production, and adaptation to various environmental conditions (see Figure 19.2).
4. Their degree of solubility:
a. Water-soluble/hydrophilic hormones: The poly- peptide/glycoprotein hormones are soluble in water and hence do not require carrier proteins for their transport in blood. In addition, they cannot freely pass through the lipid bilayer membrane and require membrane-bound receptors or transporter molecules. E.g. LH, catecholamines, GH, etc.
b. Water-insoluble/lipophilic hormones: Hormones derived from cholesterol, fatty acids, and the thyroid gland are insoluble in water and require carrier proteins for their transport in the systemic circulation.
These hormones possess intracellular (cytosolic/nuclear) receptors as they can freely cross the cell membrane. E.g. T3/T4, testosterone, aldosterone, etc.19.1.4 Synthesis of Hormones
The regulation of hormone synthesis relies heavily on fluctuations in circulating hormone or metabolite levels, as well as sensory input. Furthermore, hormones released by the hypothalamus can either stimulate or inhibit the synthesis of other hormones. When prompted by an appropriate stimulus, genes responsible for encoding specific hormones undergo increased transcription and translation. However, the precise processes of biosynthesis and post-translational modifications differ across various types of hormones, including peptides/proteins and steroids.
1. Protein or peptide hormones:
During translation, the rough endoplasmic reticulum (RER) serves as the primary site for synthesizing large precursor proteins known as “pre-prohormones” for various peptide and
FIGURE 19.2 Functions of hormones in body.
protein hormones. Within the RER, signal peptidases cleave the signal peptide from pre-prohor- mones, forming prohormones. These prohormones undergo further processing by site-specific endopeptidases, or prohormone convertases, within the Golgi apparatus (GA), leading to the production of mature hormones. These mature hormones are subsequently stored within cytoplasmic vesicles. Additionally, enzymatic cleavage, facilitated by carboxypeptidase and aminopeptidase, occurs within secretory vesicles, alongside post-translational modifications, enhancing the biological activity of specific hormones. In response to specific stimuli, such as increased intracellular calcium (Ca+2) and cAMP concentration, storage vesicles undergo fusion with the cell membrane via exocytosis, releasing hormones into the extracellular fluid.
2. Steroid hormones:
Cholesterol, either obtained from the circulation or synthesized in the smooth endoplasmic reticulum (SER) by condensing acetyl-CoA with the assistance of the rate-limiting enzyme HMG-CoA- reductase, serves as the precursor for all steroid hormones.
Cells with the capacity to release steroid hormones contain a lot of smooth endoplasmic reticulum and, as a result, lipid droplets in the cytosol. Frequently, trophic peptide hormones enhance steroid hormone-producing cells by boosting cholesterol uptake, new synthesis, and activated enzymatic machinery. A suitable stimulus activates StAR and causes a quick steroidogenic response by moving cholesterol swiftly from the OMM to the IMM. Steroidogenesis begins with cholesterol being converted into pregnenolone by an enzyme called cholesterol side chain cleavage enzyme, also known as P450scc or CYP11A1. The previous step is seen as the step that limits the rate for all types of steroid hormones. The transformation of pregnenolone into various steroid hormones relies solely on the specific cell’s mitochondrial enzymes and SER.3. General mechanism of action:
Hormones create a hormone-receptor complex (HRC) by attaching to their specific receptor on target cells or tissues. The receptors’ cellular location changes depending on the type of hormone. Peptide hormones bind to receptors on the cell membrane, while steroid hormones have receptors in both the cytosol and nucleus. Thyroid hormones have a distinctive quality in that they attach to nuclear receptors to produce their biological impacts. The HRC starts the cellular signaling pathways downstream, which ultimately regulate the metabolic and transcriptional activities of the cell either directly or indirectly.
19.1.4.1 Functioning Process of Peptide Hormones
The fact that peptide and protein hormones are attracted to water requires the existence of receptors bound to the membrane. They interact with target cells by attaching to either G-protein-coupled receptors (GPCRs) or receptor tyrosine kinases (RTK).
1. Signaling mechanism of peptide hormones:
Polypeptide and protein hormones predominantly initiate signal transduction by binding to membrane-bound G protein-coupled receptors (GPCRs). These receptors consist of three distinct regions: an extracellular domain (N-terminus), a transmembrane domain, and an intracellular domain (C-terminus).
Inactive GPCRs are associated with a GDP molecule bound to the α subunit of the G-protein. Hormone-receptor complex (HRC) formation activates GPCRs, leading to the phosphorylation of GDP to GTP and the subsequent dissociation of the GTP-bound α subunit (Ga-GTP). Ga-GTP can then activate or inhibit membrane-bound enzymes such as adenylate cyclase (AC), guanylate cyclase (GC), phospholipase C (PLC), or ion channels. G-proteins that activate specific membrane-bound enzymes or ion channels are termed stimulatory G-proteins (Gs, Gq), while those with inhibitory effects are known as inhibitory G-proteins (Gi). Receptor tyrosine kinases (RTK) are transmembrane receptors that includes tyrosine kinase receptors and tyrosine kinase-associated receptors.Both tyrosine kinase receptors and tyrosine kinase-associated receptors play crucial roles in cellular signaling. Tyrosine kinase receptors, upon hormone binding, undergo dimerization and activate their intracellular kinase domain, leading to phosphorylation of tyrosine residues and subsequent activation of downstream signaling pathways such as Ras-MAPK and Ras/PI3K/AKT. Examples include insulin acting via tyrosine kinase receptors. Tyrosine kinase- associated receptors lack intrinsic kinase domains but are coupled with protein tyrosine kinases (PTKs), notably from the Janus kinase (JAK) family. Hormone binding induces receptor dimerization, activating associated JAKs, which phosphorylate tyrosine residues on the receptor. This trigger signaling cascades such as the JAK-STAT pathway, influencing gene transcription and cellular functions. Notably, these pathways are crucial for various physiological processes, including metabolism, proliferation, and differentiation.
19.1.4.2 Functioning Process of Steroid Hormones
The majority of steroid hormone receptors are located in the cytoplasm, although some, like estrogen (E2), have receptors localized in the nucleus. These intracellular receptors feature three primary domains: the ligand-binding domain (LBD) at the C-terminus, the DNA-binding domain (DBD), and the amino-terminal domain (NTD). The LBD, housing the activation function 2 (AF2) region, serves as the binding site for steroid hormones. The DBD allows them to bind to specific regions of the genome known as hormone response elements (HRE) or steroid response elements (SRE), directly modulating the transcriptional rate of specific genes. The NTD contains the activation function 1 (AF1) region, determining optimal transcriptional activity. In their inactive state, cytosolic steroid receptors are bound with heat shock proteins (HSP). Upon binding to a steroid hormone, HSP removal and receptor phosphorylation occur, facilitating the migration and binding of hormone-receptor complexes (HRC) to SRE/HRE regions in the DNA, thus contributing to the classic steroid hormone signaling pathway. Alternatively, AF2 can indirectly regulate cellular transcription by binding to DNA-bound transcription factors like AP1 or SP1, instead of directly binding to DNA.
19.1.4 Hypothalamus
The hypothalamus is a central regulator of various bodily functions such as temperature, appetite, and emotional states, and it secretes hormones that integrate the nervous and endocrine systems. These hormones are synthesized by specialized neuroendocrine cells organized in distinct nuclei, including magnocellular and parvocellular neurons. The hypothalamic-hypophyseal portal system facilitates communication between the hypothalamus and the anterior pituitary gland, allowing for the regulation of hormone release. Hypothalamic hormones act on target cells in the pituitary gland via G protein-coupled receptors, influencing the synthesis and secretion of pituitary hormones. Pituitary tropic hormones and target hormones, originating from systemic circulation, can enter the brain and modulate hypothalamic hormone secretion directly or indirectly.
19.1.5 Pituitary
The pituitary gland, often termed the “master gland,” originates from the ectoderm and is regulated by hypothalamic stimuli to secrete hormones. Comprising the anterior, intermediate, and posterior lobes, it plays a pivotal role in regulating growth, metabolism, and reproduction through the secretion of tropic hormones. The anterior lobe, housing neuroendocrine cells responsible for synthesizing major tropic hormones, can be broadly classified into chromophils and chromophobes based on histological staining properties. Chromophils, further categorized as acidophils or basophils, actively secrete hormones, while chromophobes consist of reserve cells and undifferentiated stem cells.
19.1.6 Growth Hormone (GH)
Growth Hormone (GH), also known as Somatotropin, plays a crucial role in animal growth, spanning pre-natal and post-natal stages and varying across physiological states such as pre-pubertal, pubertal, postpubertal, and senescence. Synthesized by somatotropes, GH is initially produced as a prohormone with 217 amino acids, eventually maturing into a 191-amino acid single-chain protein with two intramolecular disulfide bridges. Binding to specific membrane-bound growth hormone receptors (GHRs), GH initiates signal transduction primarily via the JAK-STAT pathway, influencing cellular genetic machinery, and activating other pathways like Ras/MAPK and PI3K.
GH regulates body metabolism and affects various organs, including the liver, bones, skeletal muscle, adipose tissue, and gonads. A key effect of GH is stimulating hepatocytes to produce somatomedins (IGFs), particularly IGF-1 and IGF-2, which act as vital mediators of GH actions outside the liver, influencing metabolic and functional changes in target organs. GH also enhances hepatic glucose production, triglyceride secretion, and lipogenesis, while decreasing glucose utilization by skeletal muscle and adipose tissues, leading to insulin resistance and increased circulating glucose levels.
Additionally, GH stimulates insulin synthesis and secretion by pancreatic β-cells to counteract insulin resistance. Furthermore, GH promotes transcription, translation, and amino acid uptake, favoring protein anabolism and suppressing amino acid catabolism, particularly in skeletal muscles, thereby contributing to growth and development.
GH stimulates hepatocytes and adipocytes, increasing FFA, TG, and cholesterol levels for energy use, spares carbs and amino acids. It promotes protein accumulation, muscle hypertrophy via GH-IGF1 axis, and bone growth via chondrogenesis. GH induces cardiomyocyte hypertrophy, vascular expansion, and regulates testicular function. It triggers lipolysis, depleting fat reserves. Hypothalamic hormones regulate GH secretion, while IGF-1 inhibits it. Various factors like exercise and stress can stimulate GH secretion.
19.1.8 Prolactin (PRL)
Prolactin (PRL) is a multifunctional hormone with diverse roles beyond lactation. It acts through its receptors, PRLR, initiating various signaling pathways like JAK-STAT and Ras/Raf/MAPK. While primarily known for its role in lactation, PRL also influences behaviors such as maternal care and reproductive functions in animals. In mammals, PRL regulates mammary gland development, promoting lobuloalveolar growth during gestation and lactation. It stimulates the transcription and translation of casein genes, essential for milk production, and activates lactose synthesis through α-lactalbumin. This cascade of events facilitates lactogenesis and galactopoietic, ensuring adequate milk production to nourish offspring. Moreover, PRL impacts maternal behaviors, such as nest-building, nursing, and cleaning of young ones, which are crucial for offspring survival. Elevated PRL levels post-parturition contributes to the suppression of the estrous cycle, allowing the mother to focus on nurturing her offspring. Additionally, PRL secretion is influenced by estrogen levels during the estrous cycle, affecting sexual receptivity in female animals. In avian species, PRL regulates plumage patterns and influences broodiness in hens. It also stimulates the secretion of crop milk, a specialized form of nourishment provided by some bird species to their young. The regulation of PRL secretion in birds involves both dopaminergic inhibition and positive regulation by vasoactive intestinal polypeptide (VIP). Regular milking practices can also affect PRL levels, highlighting its dynamic regulation in response to environmental and physiological cues. Overall, PRL plays a pivotal role in coordinating various physiological processes essential for reproductive success and offspring care across a wide range of animal species.
19.1.9 Adrenocorticotropic Hormone (ACTH)
Adrenocorticotropic hormone (ACTH) orchestrates the synthesis of corticosteroids, vital for managing stress responses in animals. Its action is diverse, primarily focusing on stimulating the secretion of glucocorticoids to mitigate the impacts of various stressors.
Hypothalamic corticotropin-releasing hormone (CRH) triggers the synthesis of pro-opiomelanocortin (POMC) by corticotropes in the anterior lobe and neuroendocrine cells in the intermediate lobe. Cleavage of POMC in these lobes yields ACTH, a critical peptide hormone consisting of 39 amino acids. ACTH binds to the melanocortin 2 receptor (MC2-R) on adrenal cortical cells, initiating a cascade of events involving cyclic adenosine monophosphate (cAMP) production and subsequent activation of protein kinase A (PKA). This activation sets off downstream cellular pathways crucial for corticosteroid synthesis.
Upon PKA activation, there is a robust steroidogenic response marked by enhanced cholesterol synthesis, facilitated by the activation of hormone-sensitive lipase (HSL), steroidogenic acute regulatory protein (StAR), and cytochrome P450 side-chain cleavage enzyme (CYP11A1 or p450scc). This collaborative action leads to escalated conversion of cholesterol into pregnenolone, a pivotal step in corticosteroid biosynthesis. Additionally, zone-specific hydroxylases are stimulated, further diversifying corticosteroid classes.
While CRH positively regulates ACTH levels, corticosteroids, notably cortisol, exert a negative feedback loop on the CRH-ACTH axis, maintaining hormonal equilibrium essential for homeostasis. This intricate regulatory network ensures precise control over stress response mechanisms in animals.
19.1.10 Thyroid Stimulating Hormone (TSH)
Thyroid-stimulating hormone (TSH) orchestrates a finely tuned process within the thyroid gland, ensuring the synthesis and secretion of thyroid hormones align with the body’s metabolic demands. As a heterodimeric glycoprotein, TSH’s α and β chains bind specifically to thyroidstimulating hormone receptors (TSHR) on the surface of thyroid follicular cells. This interaction triggers a cascade of intracellular events, including the activation of adenylyl cyclase and subsequent elevation of cyclic adenosine monophosphate (cAMP) levels. The rise in cAMP initiates downstream signaling pathways, prominently involving protein kinase A (PKA), which in turn modulates gene transcription and translation pivotal for thyroid hormone synthesis.
TSH’s influence spans the entirety of the thyroid hormone production process. It promotes the synthesis of essential proteins such as sodium-iodide symporter protein (NIS), thyroid peroxidase, and thyroglobulin, facilitating the uptake of iodide ions and their incorporation into thyroglobulin molecules. Moreover, TSH enhances the endo- cytosis of stored colloid, thereby increasing the availability of iodinated thyroglobulin for subsequent hormone release.
Crucially, the regulation of TSH secretion is tightly controlled by feedback mechanisms. Thyrotropin-releasing hormone (TRH) and growth hormone (GH) act as primary stimulators, promoting TSH release from the anterior pituitary gland. However, the system is delicately balanced, as elevated levels of circulating thyroid hormones, particularly T3 and T4, exert negative feedback, inhibiting further TSH secretion. This intricate feedback loop ensures that thyroid hormone levels remain within a narrow physiological range, vital for metabolic homeostasis (see Figure 19.3).
Overall, TSH serves as a master regulator of thyroid function, modulating hormone synthesis and secretion to meet the dynamic metabolic needs of the organism. Its finely tuned control mechanisms underscore its critical role in maintaining metabolic balance and overall physiological health.
The thyroid gland, a focal point in physiological psychology discussions, is a roughly 30-gram organ distinguished by its vivid red hue and encased in connective tissue. Comprising two lobes joined by an isthmus, it originates from a pharynx floor invagination during fetal development’s second month, gradually releasing hormones with profound physiological impacts. This article explores the thyroid gland’s pivotal role and the significance of its hormones.
Measuring around 5 centimeters in length and 2 centimeters in width and thickness, the thyroid lobes are relatively large endocrine glands nestled alongside the trachea, larynx, and carotid artery sheath. The isthmus, connecting the lobes anteriorly to the second and third tracheal rings, is crucial for structural integrity. Blood supply to the thyroid is facilitated by the superior and inferior thyroid arteries.
The thyroid tissue comprises over one million follicles or closed acini, each with a diameter of 150 to 300 microns. These follicles’ walls consist of a single layer of cuboidal epithelial cells, enclosing colloid - a substance predominantly composed of thyroglobulin, a protein synthesized by follicular cells. Parafollicular cells interspersed among the follicles produce calcitonin, a secondary hormone involved in calcium metabolism. The thyroid’s extensive vascular branching ensures efficient blood supply and hormone entry into circulation, with both sympathetic and parasympathetic nerves innervating the gland.
The thyroid gland secretes two main iodine-containing hormones - thyroxine (T4) and triiodothyronine (T3) - synthesized by follicular and parafollicular cells. Thyroxine, or T4, features four iodine atoms, while triiodothyronine, or T3, contains three. Follicular cells synthesize thyroglobulin from blood-borne tyrosine, storing it within follicles’ lumen. These follicles, grouped into lobules, actively participate in hormone synthesis.
Approximately 93% of thyroid hormone secretion comprises thyroxine, with the remaining 7% being triiodothyronine. However, tissues predominantly utilize triiodothyronine, derived from thyroxine conversion. Thyroglobulin iodination, facilitated by follicular cells, progresses from monoiodotyrosine to diiodotyrosine, ultimately forming triiodothyronine and thyroxine. Upon synthesis, these iodine-containing hormones bind to plasma proteins for transportation to target tissues, where they modulate gene transcription, enhance metabolic activity, and influence growth and development.
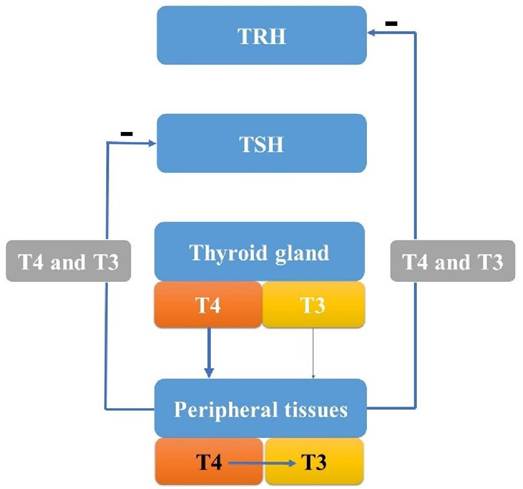
FIGURE 19.3 Thyroid hormone: function and mechanism of action.
Thyroid hormones play diverse physiological roles, including stimulating carbohydrate and fat metabolism, regulating plasma lipid levels, increasing basal metabolism, and influencing cardiac function, nervous system activity, and muscle function. Moreover, thyroid hormone imbalances can affect sleep patterns, muscle tremors, and the secretion of other hormones.
19.1.11 Pituitary Gonadotropins (LN and ESH)
Pituitary gonadotropins play a central role in regulating reproductive functions, ensuring the production of steroid hormones and the maturation of gametes, particularly in animals after reaching sexual maturity, which is essential for maintaining optimal reproduction in farm animals.
Comprising α and β subunits, gonadotropins are glycoprotein heterodimers, with the unique composition of the β subunit determining their specific activities. The glycosylation of specific residues within these subunits is crucial, influencing their effects and duration within the body. Upon binding to their respective receptors, such as the luteinizing hormone receptor (LHR) and follicle-stimulating hormone receptor (FSHR), gonadotropins initiate intricate signaling cascades. These cascades lead to an increase in cyclic adenosine monophosphate (cAMP) levels and activate protein kinase A (PKA), which subsequently regulates various cellular processes essential for steroid hormone production and cell cycle control.
Luteinizing hormone (LH) acts on thecal cells within ovarian follicles, stimulating the production of testosterone, a precursor for estrogen synthesis and a key factor in ovulation in females. In males, LH targets Leydig cells, promoting testosterone production, which is vital for male fertility. Furthermore, LH facilitates the transformation of granulosa cells into luteal cells and plays a pivotal role in regulating male and female reproductive processes.
Follicle-stimulating hormone (FSH) predominantly affects granulosa cells in females, fostering estrogen synthesis and supporting spermatogenesis and libido maintenance in males by acting on Sertoli cells. FSH sensitivity is highest in the dominant follicle in females and plays a critical role in regulating essential processes in males.
Gonadotropin-releasing hormone (GnRH) serves as the primary regulator of gonadotropin secretion, orchestrating the intricate hormonal balance necessary for reproductive health. Additionally, factors such as estrogen, activin, and inhibin modulate FSH and LH secretion, contributing to the intricate regulation of reproductive processes to ensure optimal fertility in animals.
19.1.12 Posterior Pituitary
The posterior pituitary, originating from neural ectoderm, comprises axons stemming from magnocellular (MC) neurons found in the paraventricular nucleus (PVN) and supraoptic nucleus (SON) of the hypothalamus. Upon receiving appropriate stimulation, these nerve endings in the posterior pituitary release oxytocin (OT) and antidiuretic hormone (ADH). Additionally, the neurohypophysis contains glial cells known as pituicytes.
Both oxytocin and ADH are nonapeptides, differing slightly in their amino acid composition. They are synthesized within MC neurons, accompanied by their specific precursor transport protein called neurophysin, and stored as secretory granules. However, during axonal transport, the transport protein component is cleaved to yield the active hormone. Action potentials generated in response to neural stimuli on MC neurons trigger the release of these hormones from their nerve terminals (see Figure 19.4).
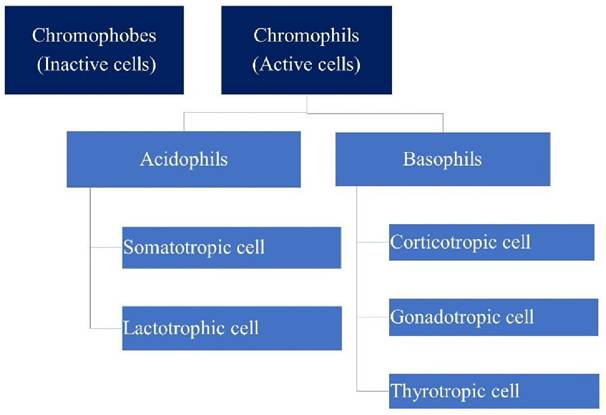
FIGURE 19.4 Cell types in the pituitary gland.
19.1.13 Oxytocin
Oxytocin, well-known for its crucial role in childbirth, also affects milk ejection, sperm transport, social bonding, and ovulation in animals. It binds to oxytocin receptors (OTR), specific G protein-coupled receptors, triggering the phospholipase C (PLC) system to generate secondary messengers such as diacylglycerol (DAG) and inositol triphosphate (IP3). These secondary messengers activate protein kinase C (PKC) and release calcium ions (Ca+2) from the endoplasmic reticulum. Elevated Ca+2 levels activate myosin-light chain kinase (MLCK), while PKC inhibition of myosin- light chain phosphatase (MLCP) increases actin-myosin bridge formation, initiating smooth muscle contraction.
The afferent neural stimuli resulting from fetal entry into the cervical region induce oxytocin release, triggering Ferguson’s reflex, a neuroendocrine reflex. This reflex stimulates smooth muscle cells in the myometrium to initiate uterine contractions, facilitating fetal expulsion, hence its moniker, the “birth hormone.” Oxytocin is also released during and after mating in both male and female animals, aiding spermatozoa transport in the reproductive tracts of both sexes.
Contraction of myoepithelial cells surrounding the alveoli prompts milk ejection from the mammary gland. In dairy animals, tactile stimulation of the udder, along with visual or auditory stimuli, induces oxytocin secretion, facilitating milk ejection.
Oxytocin additionally stimulates prostaglandin F2α (PGF2α) synthesis, which plays a crucial role in follicular membrane rupture, leading to ovulation.
19.1.14 Anti-Diuretic Hormone
ADH, crucial for water conservation during dehydration, combats urine water loss and prevents ECF osmolality rise.
It binds to vasopressin receptors (VR), influencing various tissues depending on receptor type: V1, V2, and V3.
Binding to V2 receptors in tubular epithelial cells triggers cAMP elevation, prompting aquaporin gene activity. This leads to aquaporin insertion, enhancing water reabsorption and displaying ADH’s anti-diuretic effect.
V1 receptor activation in arteriolar smooth muscle cells stimulates PLC, generating DAG and IP3. This leads to calcium influx, inducing muscle contraction, hence its alias, vasopressin. This constriction reduces GFR, conserving water and maintaining blood pressure during dehydration.
Osmoreceptors in the SON detect ECF osmolality shifts, stimulating ADH release. Heart stretch receptors also prompt ADH secretion in response to blood volume reductions (see Figure 19.5).
19.1.15 Pineal Gland
Often termed the “biological clock,” the pineal gland regulates mammalian circadian rhythms by producing melatonin and comprises glial cells and pinealocytes.
Melatonin, derived from tryptophan, undergoes hydroxylation and carboxylation to form serotonin and then N-acetyl serotonin, respectively, through enzymatic processes involving AANAT and ASMT.
While the pineal gland is essential for circadian rhythm control, the suprachiasmatic nucleus (SCN) of the hypothalamus houses the underlying cellular and molecular mechanisms. SCN’s sympathetic innervation activates pineal gland melatonin synthesis during darkness by stimulating AANAT and ASMT gene expression. Additionally, rhythmic clock gene expression, particularly Perl, contributes to melatonin synthesis, peaking at night. Melatonin then influences clock gene activity in the hypothalamus and peripheral tissues.
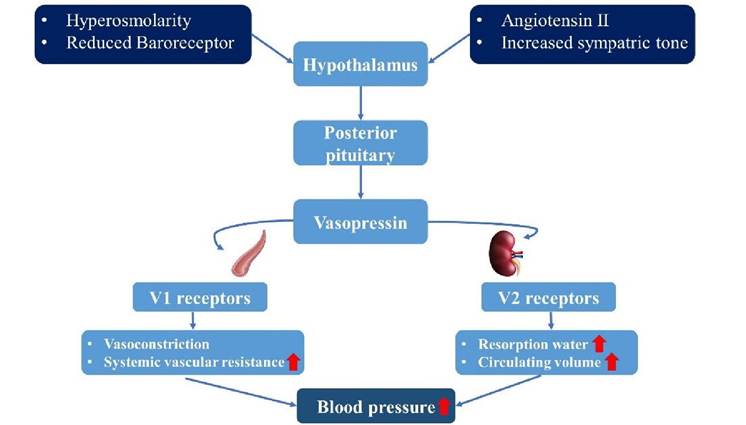
FIGURE 19.5 Mechanism of secretion and action of ADH.
TABLE 19.1
Thymic Hormones and their Function Concerning the Immune System
| Thymic Hormone Chemical Nature | Function in the Immune System |
| Thymosin α1 Peptide Thymosin β4 Peptide Thymulin Peptide | Stimulates T cell development, enhances T cell function, modulates immune responses Promotes wound healing, regulates immune cell migration and differentiation Stimulates T cell maturation and differentiation, enhances T cell function, modulates immune responses |
| Thymopoietin Peptide | Supports T cell development and maturation, regulates immune cell proliferation |
Melatonin receptors, including MT1 and MT2, inhibit adenylyl cyclase activity, reducing cAMP levels, and affecting various signaling pathways.
While known for its circadian rhythm, sleep, and reproductive regulation, melatonin also combats oxidative stress and modulates the cell cycle and apoptosis. Its anti-apoptotic effects are crucial for follicle growth, oocyte maturation, and luteinization, ensuring fertility. In males, melatonin’s antioxidative properties protect spermatozoa, supporting fertility by impacting steroidogenesis and mitigating oxidative stress on gametes.
Melatonin levels fluctuate with seasons, influencing breeding cycles by signaling optimal reproductive timing. In short-day breeders like sheep, increased melatonin levels with shorter daylight durations trigger the estrous cycle onset, while in long-day breeders, it aligns with longer daylight periods. Melatonin’s effects on seasonal breeding alter pituitary gonadotropin and thyrotropin secretion patterns.
19.1.16 Gastrointestinal Hormones
Considered as the only site for nutrient assimilation, the gastrointestinal tract (GIT) also produces hormones that are responsible for regulating the secretions of exocrine glands, gastrointestinal motility, cellular proliferation, and differentiation in the GIT. The first hormone to be discovered by Bayliss and Starling (1902) is secretin, which is a gut hormone. Considered the largest endocrine organ in the animal’s body, the GIT produces more than 20 peptide hormones.
The hormone-producing cells are also known as enteroendocrine cells (EECs), interspersed between gut mucosal cells in the GIT. They are derived from the pluripotent intestinal stem cells in the intestinal crypts. Different types of EECs are present in the GIT, with each cell type having the ability to synthesize and secrete at least one kind of hormone. The hormones thus produced can also act as autocrine or paracrine or neurocrine factors on the nearby target cells. Based on structural homologies, the gut hormones are further classified into different families: the secretin family (secretin, glucagon-like peptides, VIP), the gastrin family (gastrin, CCK), and the PP-fold family (Neuropeptide Y (NPY), PYY, pancreatic polypeptide).
The gut hormones have the unique property of activating at least one specific type of GPCR present on the target tissues (Table 19.1). The signal transduction mechanisms include the activation of adenylyl cyclases, protein kinases, and membrane-bound ion channels. With overlapping target organs, multiple gut hormones are needed to produce the desired biological effects. The major biological effects of gut hormones include the regulation of ingestion, digestion, and the metabolism of nutrients (see Table 19.2).
19.1.17 Pancreas
The pancreas, often termed the “abdominal gland,” is an indispensable component of the digestive system, orchestrating a symphony of functions crucial for metabolic equilibrium. Positioned anteriorly to the lumbar vertebrae, its name, rooted in Greek, underscores its multifaceted role. With its distinct pinkish-gray hue and robust texture, spanning about 15 to 20 centimeters in length, 2.5 to 4 centimeters in width, and weighing approximately 90 grams, the pancreas commands attention.
Within the realm of digestion, the pancreas plays a pivotal role, acting as a hub for the production of digestive juices and enzymes crucial for breaking down complex nutrients such as carbohydrates, fats, and starches. Simultaneously, it operates as an endocrine powerhouse, secreting hormones that meticulously regulate blood sugar levels and stomach acid secretion. Deep within its tissue lie specialized cell clusters known as pancreatic islets, each contributing unique hormones: beta cells produce insulin, alpha cells yield glucagon, delta cells release somatostatin, and PP cells generate pancreatic polypeptides.
Functioning as an exocrine gland, the pancreas diligently synthesizes an array of digestive enzymes, including trypsin, chymotrypsin, amylase, and lipase, essential for efficient nutrient breakdown. These enzymes embark on their digestive journey through the pancreatic duct, converging with the common bile duct to form the ampulla of Vater, culminating in their discharge into the duodenum. Meanwhile, the endocrine facet of the pancreas, embodied by the Islets of Langerhans, dispatches hormones such as insulin and glucagon directly into the bloodstream, meticulously overseeing blood sugar levels. In the grand scheme of metabolic regulation, insulin and glucagon emerge as
TABLE 19.2
Gastrointestinal-Secreted Hormones: Source, Chemical Structure, Receptors and Effects on Feed Intake
| Chemical | ||||
| Hormone | Source | Structure | Receptors | Effects on feed intake |
| Ghrelin | Stomach, Small Intestine | Peptide | Growth hormone secretagogue receptor (GHS-R) | Stimulates appetite, increases feed intake |
| Cholecystokinin (CCK) | Duodenum, Jejunum | Peptide | CCK-A receptors | Suppresses appetite, reduces feed intake |
| Glucagon-like peptide 1 (GLP-I) | L-cells of the intestine | Peptide | GLP-1 receptors | Suppresses appetite, reduces feed intake |
| Peptide YY (PYY) | L-cells of the ileum and colon | Peptide | Y2 receptors | Suppresses appetite, reduces feed intake |
| Leptin | Adipose tissue | Protein | Leptin receptors in the hypothalamus | Suppresses appetite, reduces feed intake |
| Insulin | Pancreas | Peptide | Insulin receptors | Modulates appetite, can reduce feed intake |
| Neuropeptide Y (NPY) | Hypothalamus, Enteric nervous system | Peptide | NPY receptors | Stimulates appetite, increases feed intake |
| Gastrin | Stomach, Duodenum | Peptide | Cholecystokinin B receptors | Stimulates gastric acid secretion, may influence appetite |
| Secretin | Duodenum | Peptide | Secretin receptors | Regulates bicarbonate and water secretion, may indirectly affect appetite |
| Motilin | Duodenum, Jejunum | Peptide | Motilin receptors | Stimulates gastrointestinal motility, may influence feeding behavior |
| Neurotensin | Small Intestine | Peptide | Neurotensin receptors | Regulates intestinal motility and secretion, may have an effect on appetite |
pivotal players, engaged in a delicate dance to maintain blood sugar homeostasis (see Table 19.3).
19.1.17.1 Insulin
Function: Acting as a key regulator of glucose metabolism, insulin is released by beta cells in response to elevated blood sugar levels. It facilitates glucose uptake by cells, promoting glycogen storage in the liver and muscle cells, and converting excess glucose into fat.
Storage: Insulin promotes the storage of excess glucose in the liver and muscle cells in the form of glycogen, facilitating long-term energy storage.
Protein Synthesis: Insulin facilitates protein synthesis by promoting the uptake of amino acids into cells.
Lipid Metabolism: Inhibiting fat breakdown (lipolysis) and promoting fat storage, insulin contributes to overall lipid metabolism.
Anabolic Effects: Insulin fosters growth, energy storage, and cellular repair (see Table 19.3).
19.1.17.2 Glucagon
Function: Glucagon serves as a counter-regulatory hormone to insulin, released by alpha cells when blood sugar levels drop. It stimulates glycogen breakdown (glycogenolysis) in the liver, promoting glucose release into the bloodstream.
Glycogenolysis: Activating liver enzymes, glucagon breaks down glycogen into glucose, ensuring adequate blood sugar levels.
TABLE 19.3
Pancreas Hormones: Chemical Nature, Effects, and Plasma Half-Life
| Plasma Half-life | |
| Hormone Chemical Nature | Effects (minutes) |
| Insulin Peptide | Facilitates glucose uptake into cells, promotes glycogen synthesis, ~5 inhibits gluconeogenesis and lipolysis |
| Glucagon Peptide | Stimulates glycogenolysis, gluconeogenesis, and lipolysis, raises blood ~5-10 glucose levels |
| Somatostatin Peptide | Inhibits the release of insulin and glucagon, slows down gastrointestinal ~2-3 motility and nutrient absorption |
| Pancreatic Polypeptide (PP) Peptide | Inhibits exocrine pancreatic secretion, regulates gastrointestinal function ~30 |
Gluconeogenesis: Glucagon prompts the liver to produce glucose from non-carbohydrate sources, maintaining a steady glucose supply.
Lipolysis: Glucagon promotes fat breakdown into fatty acids and glycerol, serving as an alternative energy source.
Opposing Actions: Glucagon’s actions oppose those of insulin, preventing hypoglycemia and maintaining blood glucose levels within a narrow range (see Table 19.3).
19.1.18 Reproduction Hormones
19.1.18.1 Placenta
The placenta acts as the site of attachment of the fetus, delivers nutrients and gases derived from maternal circulation, prevents the fetal allograft from the maternal immune system, and eliminates fetal metabolic-waste products. In addition, it acts as a transient endocrine organ by producing hormones that are vital for the maintenance of gestation, fetal growth, and parturition (see Table 19.4).
19.1.18.2 Progesterone
Progesterone stands as an absolute requirement for the maintenance of gestation in all mammals. Although corpus luteum is the primary source of progesterone in domestic animals, placental-derived progesterone also helps in the maintenance of gestation. The shift of progesterone production from the corpus luteum to the placenta is regarded as an essential phenomenon seen in the case of equines, sheep, and primates. This luteo-placental shift in progesterone synthesis has minor relevance in goats and pigs. Luteal-progesterone plays a prominent role in myometrial quiescence, endometrial growth and differentiation, immunosuppression to prevent fetal rejection, and cervical closure. However, the progesterone derived from the placenta is speculated to be useful for species with longer gestation (see Table 19.4).
19.1.18.3 Estrogen
Unlike follicle that produces 17β-estradiol, placenta in ungulate animals primarily secretes estrone in sulfate form. In primates, placental estrogen is implicated in trophoblast differentiation, mammary gland development, and uteroplacental blood flow. However, in cattle, maternal estrogen levels rise along with a concomitant decrease in progesterone levels at the end of gestation period due to an increased conversion of progesterone to estrogen by CYP17A1 (17- α-hydroxylase) in the placenta. Thus, the increased circulatory levels of estrogen lead to the ductular development of the mammary gland, remove the progesterone-mediated negative feedback on lactation, excite the myometrial tissue, and prepare the birth canal for parturition.
19.1.18.4 ChorionicGonadotropins
As of now, only two chorionic gonadotropins: equine chorionic gonadotropin (eCG) and human chorionic gonadotropin (hCG) are reported in equines and primates, respectively. Like the pituitary gonadotropins (LH & FSH), the chorionic gonadotropins are made of a common α-glycoprotein chain. However, β-chains of chorionic gonadotropins (βCG) are similar to the βLH chain, except that they are heavily glycosylated. The hCG released by blastocyst in primates is responsible for the maternal recognition of pregnancy and maintenance of corpus luteum by binding to LH receptors. Whereas, distinct areas of the placenta known as endometrial cups secrete eCG to promote the formation of accessory corpora lutea, thereby helping in the maintenance of gestation.
19.1.18.5 Placental Lactogen (Chorionic Somatomammotropin)
The binucleate cells of trophectoderm in ruminants, primates, and rodents secrete placental lactogen, a hormone that has both somatotropic and lactogenic functions. Mature placental lactogen (PL) is a single polypeptide chain with 200 amino acids. In bovines, PL shares a structural homology of 50% and 23% with PRL and GH, respectively. The circulatory levels of PL start raising from Day 30 onwards, reaching a peak during the last trimester, and begin to fall when the animal approaches parturition. In ruminants, PL has luteotrophic actions and augments progesterone secretion from the corpus luteum. It plays a major role in the partitioning of maternal nutrients to support fetal growth by stimulating the uptake of maternal nutrients, and glycogenesis in fetal tissues. In addition, it stimulates lobuloalveolar growth in the mammary gland and exerts galactopoietic effects by stimulating dry matter intake.
19.1.18.6 Relaxins
They are peptide hormones belonging to the insulin family, produced primarily by the corpus luteum and placenta. Relaxin1 (RLN 1) plays a major role in reproductive functions in mammals. It is a polypeptide heterodimer hormone with 53 amino acids. The production of RLN1 rises at the end of gestation to promote cervical ripening, dilatation of pubic symphysis, and relaxation of the sacrosciatic ligament to aid the process of parturition.
19.1.19 Ovary and Testes
Gonads refer to the testes and ovaries present in male and female animals, respectively. They are responsible for the production of gametes and hormones, which are crucial elements governing sexual reproduction in animals (Table 19.4). The hypothalamic-pituitary axis secretes GnRH, LH, and FSH, which are essential for regulating the endocrine and gametogenic activities of gonads. In turn, the gonadal hormones produced exert negative inhibition on the release of hypothalamic GnRH and pituitary gonadotropins (see Table 19.4).
TABLE 19.4
Reproductive Hormones: Source, Chemical Structure, Function, and Characteristics
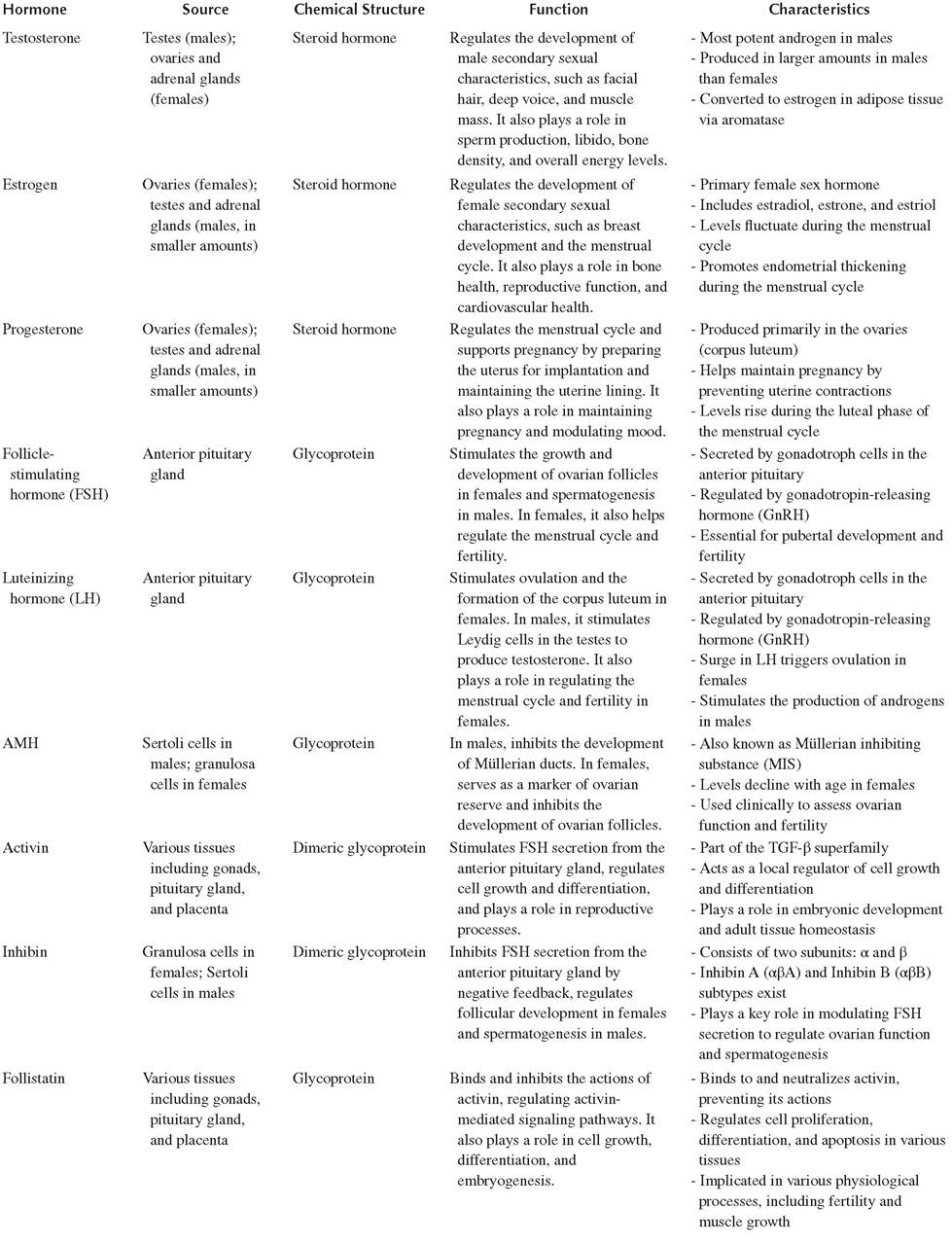
19.1.20 Other
19.1.20.1 The Heart as an Endocrine Organ
The endocrine activity of the heart comprises the release of various natriuretic peptide hormones from the cardiomyocytes. The atrial natriuretic peptide family includes three structurally similar peptides, i.e., atrial natriuretic peptide (ANP), brain or B-type natriuretic peptide (BNP), and C-type natriuretic peptide (CNP). The secretion of these hormones enables the heart to participate in regulating the circulatory volume and systemic blood pressure in animals.
The target effects of natriuretic peptides are mediated by cell membrane-bound receptors coupled with guanylyl cyclase. Three different types of receptors were identified: NPR-A, NPR-B, and NPR-C. ANP and BNP exert their biological effects by binding to NPR-A. The binding of ANP or BNP to NPR-A results in a conformational change with consequent activation of the intracellular guanylyl cyclase domain. The cGMP formed from GTP in the presence of guanylyl cyclase is regarded as the secondary messenger in the signal transduction mechanism. The elevated cGMP levels affect various cGMP-dependent protein kinases (PKGs), cGMP gated ion channels, and cGMP-regulated phosphodiesterases to bring about specific target effects. Kidneys, blood vessels, and the adrenal cortex are regarded as the principal target tissues of natriuretic peptides.
The natriuretic peptides (except CNP) are responsible for lowering the blood volume by enhancing the renal excretion of water (diuresis) and sodium (natriuresis). Apart from kidneys, they also cause the relaxation of vascular smooth muscles thereby bringing down the arterial pressure. They act on the pituitary gland, adrenal cortex, and small intestine to inhibit ADH, aldosterone secretion, and sodium absorption, respectively.
19.1.20.2 The Kidney and the Liver as Endocrine Organs
Erythropoietin (EPO) is a glycoprotein hormone (167 amino acids) released from the kidneys in response to hypoxia. The fetal liver and kidneys are major sites for the production of EPO. The interstitial peritubular cells in proximal tubules synthesize and secrete EPO during hypoxia and anemic stress.
The activation of erythropoietin receptors (EPOR) when bound with EPO results in their dimerization and phosphorylation by Janus kinase 2 (JAK2). The transphosphorylation of EPOR initiates the phosphorylation of signal transducer and activator of transcription 5 (STAT5), PI3K/AKT, and SHC/MAPK pathways to regulate cellular proliferation and survival. The EPOR is distributed majorly in erythrocyte progenitor cells residing in the bone marrow, thus making them the primary target tissue.
EPO is essential for the survivability of erythrocyte progenitor cells and their subsequent differentiation to colonyforming unit-erythrocyte (CFU-E) cells. It results in the swift passage of erythroblasts through different stages in erythropoiesis, thereby increasing the rate of erythropoiesis (Figure 19.2). Apart from the erythrocyte progenitors, EPO is found to have angiogenic effects on the endothelial cells and neurotrophic effects in the brain and regulate bone mass and formation of skeletal muscle fibers.