Introduction to regions
The anatomical descriptions are based primarily on canine anatomy but they are also relevant to most domestic animals.
There are three main components to the nervous system: the central nervous system (CNS), the peripheral nervous system (PNS) and the autonomic nervous system (ANS).
Peripheral nervous system
Key points
■ The peripheral nervous system consists of the nerves and ganglia located outside the brain and spinal cord.
■ The myelin sheath surrounding the peripheral axons is formed by Schwann cells, whereas CNS axons are myelinated by oligodendrocytes.
■ Dorsal and ventral nerve roots attach on each side of the spinal cord and carry sensory and motor axons, respectively. Lateral to the cord, the dorsal and ventral roots fuse to form mixed, spinal nerves. Adjacent spinal nerves may fuse, in a plexus, to form named nerves in the periphery.
■ There are 12 pairs of cranial nerves that innervate the head and extend into the body.
■ Areas of sensory innervation of the skin are categorised as dermatomes, cutaneous zones and autonomous zones.
■ Somatic lower motor neurons innervating striated muscle of the body have their cell bodies sited in the CNS; their axons travel in the PNS to connect to the muscle at the neuromuscular junction. A motor unit comprises a single lower motor neuron (LMN) and the group of muscle fibres it innervates.
The PNS consists of the nerves and ganglia located outside the brain and the spinal cord and principally functions to connect the central nervous system (CNS) to the head, body, limbs and viscera. With respect to nomenclature, a ‘nerve(s)’ is by definition, in the PNS, making the word ‘peripheral’ (as in ‘peripheral nerve’) redundant; it is myelinated by Schwann cells. Unlike the CNS, the PNS is not protected by bone, leaving it more vulnerable to mechanical injury.
Schwann cells form the insulating myelin sheaths surrounding peripheral axons, whereas in the CNS that task is performed by oligodendrocytes. The change from oligodendrocytes to Schwann cells occurs where the dura mater surrounding the spinal cord abuts the perineurium at the origin of the spinal nerves (Fig. 1.1). Afferent and efferent axons of the PNS form the spinal and cranial nerves (CNN).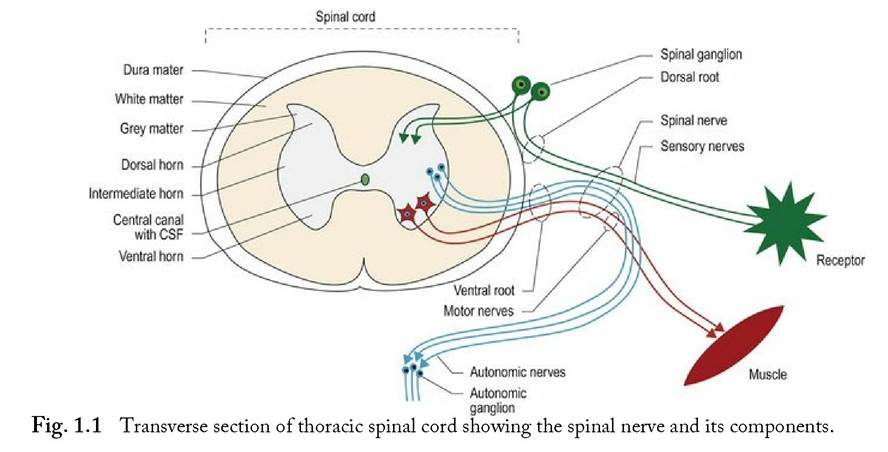
Spinal nerves
Spinal nerves arise as roots from the spinal cord. A dorsal and a ventral root attach on each side of the spinal cord, and define each spinal cord segment. For example, the third cervical spinal cord segment has two dorsal roots and two ventral roots attaching to it. The dorsal roots convey primarily sensory nerve fibres into the spinal cord. Each dorsal root contains a spinal ganglion (old name ‘dorsal root ganglion’), housing the nerve cell bodies of these sensory fibres (Fig. 1.1). The ventral roots convey motor nerve fibres away from the spinal cord. Motor fibres may be somatic and innervate striated muscle, or autonomic and innervate smooth or cardiac muscle. The dorsal and ventral roots fuse at the level of the intervertebral foramen to form a spinal nerve. Distal to the intervertebral foramen, the mixed sensory and motor spinal nerve usually splits into a dorsal and ventral branch. The dorsal branch supplies the epaxial muscles and skin, while the ventral branch supplies the hypaxial muscles and skin. A third branch, carrying autonomic fibres, may also arise and pass ventrally towards the midline to supply the viscera.
Spinal nerves may remain as single, discrete nerves all the way out into the periphery, in which case they are named for the number of the spinal cord segment from which they arise, e.g. ‘C3, ventral branch’ is the ventral division of cervical spinal nerve 3; it supplies sensory and motor innervation to the hypaxial tissue of the neck. Alternatively, in a nerve plexus, the ventral branches of two or three adjacent spinal nerves may fuse, giving rise to nerves with specific names, such as radial nerve and femoral nerve (Fig.
1.2).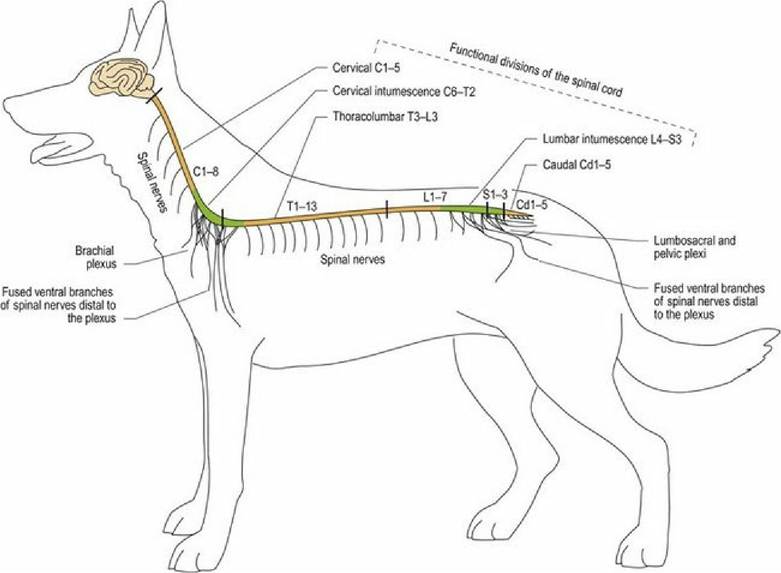
Fig. 1.2 Anatomical and functional spinal cord regions and the main nerve plexi.
A cranial nerve arises from the brain; it is known by both the number of the nerve and a specific name, e.g. trigeminal nerve or cranial nerve, CN V (Fig. 1.6A).
The PNS comprises afferent (sensory) fibres and their receptors, and efferent (motor) fibres that link to effectors, such as striated, cardiac and smooth muscle.
Afferent nerve fibres originate at different types of sensory receptors and most of their fibres only synapse once they reach the CNS. The sensory nerve cell bodies of spinal nerves are located in spinal ganglia sited at the level of the intervertebral foramen. Cranial nerve ganglia are located just near, or inside, the neurocranium. The area of skin innervated by a spinal nerve is called a dermatome, while the area of skin innervated by a specific named nerve, which originates from two or more spinal nerves (for example, the radial nerve) is called a cutaneous zone. Adjacent dermatomes and cutaneous zones usually overlap. The area of skin innervated purely by one nerve is called an autonomous zone (Fig. 1.3). Autonomous zones are found on the head
(Figure 10.13) as well as the body and limbs (Table 1.1).
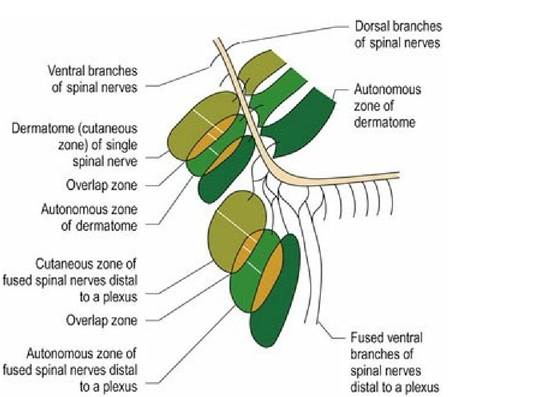
Fig. 1.3 Sensory innervation of the skin depicting the basis of dermatomes, cutaneous zones and autonomous zones. The zones in this diagram are conceptual and not anatomically accurate to the thoracic limb.
Table 1.1 Autonomous zones in the canine limb
| Thoracic limb nerves | Site for testing autonomous zone | Pelvic limb nerves | Site for testing autonomous zone |
| Musculocutaneous | Medial aspect of | Saphenous | Medial aspect of crus |
| nerve | antebrachium | nerve | (stifle to hock) |
| Radial nerve | Dorsal aspect digits 2, 3,4 | Peroneal nerve | Dorsal aspect digits 2, 3,4 |
| Ulnar nerve | Lateral aspect digit S | Tiblal nerve | Plantar aspect digits 3 and 4 |
Knowing the position of dermatomes and autonomous zones is useful for assessing sensory function of the PNS.
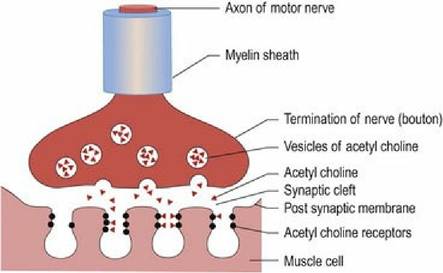
However, there are species differences, for example, the horse does not have an autonomous zone specific to the radial nerve.Motor neurons can be defined as upper motor neurons (UMNs) or lower motor neurons (LMNs). The UMN is confined to the CNS and its axon influences activity of LMNs. They are the ‘managers’ of the motor system (see Chapter 4). The LMNs are found in cranial nerves, originating from the brainstem, and spinal nerves from the spinal cord. They form synapses at the neuromuscular junction and innervate striated muscle, smooth or cardiac muscle. They are the ‘workers’ of the motor system. The neurochemical that connects the electrical activity of the motor nerve to the striated muscle is acetylcholine (ACh). A nerve impulse arriving at the nerve termination triggers release of ACh (Fig. 1.4). The ACh crosses the synaptic cleft, binds to the receptors on the post-synaptic membrane and may stimulate muscle membrane depolarisation and muscle contraction, depending on stimulus strength and amount of ACh released. The ACh is broken down by acetylcholine esterase and recycled back into the distal end of the LMN. For striated muscle, the motor unit is defined as an axon and the muscle fibres it innervates. Motor units range in size from 3-150 muscle fibres per axon for muscles with fine versus coarse control, respectively. Small motor units are found in extraocular muscles, whereas large motor units are found in the large, postural muscles, e.g. the quadriceps femoris muscle.
Fig. 1.4
The neuromuscular junction.
Cranial nerves
In domestic mammals, there are 12 pairs of cranial nerves. Like the spinal nerves, these may convey sensory, motor and autonomic fibres of the parasympathetic nervous system (see p4). Cranial nerves are abbreviated to CN (singular) or CNN (plural).
Sensory ganglia are located near, or just inside, the neurocranium whereas the somata of motor nerves are located primarily in the brainstem.
The exception is the external branch of the accessory nerve (CN XI); these fibres originate in the cervical spinal cord. The cranial nerve fibres generally remain distinct from other cranial nerves, but occasionally they may intermingle. For example, CNN X and XI fibres mix to form the recurrent laryngeal nerve. The nerves and their branches have specific names, e.g. auriculopalpebral branch of the facial nerve, CN VII.Using Roman numerals, the nerves are numbered in rostro-caudal sequence based on their attachment to the brain. Assessing the function of the cranial nerves is an integral part of the clinical examination. They are covered in more detail in Chapter 10.
Autonomic nervous system
Key points
■ The ANS consists of efferent and afferent fibres that innervate smooth and cardiac muscle of viscera. It has both central and peripheral components and its fibres travel in the PNS via cranial and spinal nerves.
■ It is divided into sympathetic (‘fight or flight’) and parasympathetic (’rest and digest’) components.
■ Anatomically, these divisions are also referred to as the thoracolumbar and craniosacral systems based on the origin of fibres from the CNS.
The ANS is variably defined depending on the source, but consists of efferent fibres that innervate smooth and cardiac muscle of viscera. In this book, afferent nerves from the viscera are considered to be part of the ANS. The ANS has both central and peripheral components; the latter are found in cranial and spinal nerves.
The ANS functions largely without conscious input to maintain homeostasis in the body at rest and during times of stress. It is divided on the basis of function into two components, the parasympathetic and sympathetic nervous systems. The parasympathetic system is known as the ‘rest and digest’ system and controls day-to-day activities such as digestion, elimination of wastes and activity that contributes to homeostasis. The sympathetic system is called the ‘fight or flight’ system and prepares the body for these activities by increasing heart and respiratory rate, dilating the pupils, redirecting blood flow to major limb muscles and away from activities such as digestion.
Anatomically, the two systems arise from different areas of the CNS. The parasympathetic system arises from the brainstem and sacral spinal cord, hence it is called the craniosacral system. The sympathetic system arises from the thoracolumbar spinal cord and is called the thoracolumbar system. For both parasympathetic and sympathetic systems, the nerve fibres leaving the CNS will synapse in a ganglion once en route to their target organ.
Central nervous system
Key points
■ The CNS consists of the spinal cord and the brain.
■ The spinal cord is divided into five clinically important, functional regions: cervical, cervical intumescence, thoracolumbar, lumbar intumescence and caudal regions.
■ Anatomically, the divisions of the brain are the forebrain, the midbrain and the hindbrain. Functionally, the divisions of the brain are the forebrain, brainstem and cerebellum and each portion is associated with distinctive functions.
■ Grey matter houses neuronal somata. White matter is formed by axons, many of which have lipid-rich myelin sheaths, causing the white appearance.
■ In the spinal cord, neurons forming the sensory, autonomic or motor regions of grey matter are centrally located; they form longitudinal columns extending the length of the cord. In the brainstem, the grey matter columns are fragmented into clusters of cells called nuclei. In the forebrain, there are both superficial and deeply located grey matter.
The CNS comprises the spinal cord and brain. Externally, it is covered by three layers of fibrous, supporting tissue, the meninges (see Chapter 3). Internally, the ventricular system forms a series of fluid-filled spaces that connect to the externally located, subarachnoid space. The ventricular system contains cerebrospinal fluid (CSF) (see Chapter 3).
Spinal cord
The spinal cord is protected by the vertebral column. It is divided into five anatomical regions based on the vertebral column, and five functional regions based on the innervation of the limbs. Table 1.2 refers to canine neuroanatomy.
Table 1.2 Anatomical and functional regions of the canine spinal cord (see also Fig. 1.2)
| Anatomical region | Spinal cord segments | Functional region and area supplied | Spinal cord segments |
| Cervical | C1-C8 | Cervical: neck | CI-CS |
| Thoracic | Tl-Tl 3 | Cervical intumescence: thoracic limb | C6-T2 |
| Lumbar | Ll-L7 | Thoracolumbar: thorax and abdomen | T3-L3 |
| Sacral | SI-S3 | Lumbar intumescence: pelvic cavity, pelvic limb, perineum | L4-S3 |
| Caudal | CdI-CdS | Caudal: tail | CdI-CdS |
The number of vertebrae varies for different types of animals; consequently the number of spinal cord segments will also vary (Table 1.3).
Table 1.3 Numbers of vertebrae in different types of animals
| Animal | Cervical | Thoracic | Lumbar | Sacral | Caudal |
| Dog and cat | 7 | 13 | 7 (occasionally 61 | 3 | 20+ |
| Horse | 7 | IB | 6 | S | 20 |
| 0κ | 7 | 13 | 6 | 5 | 18-20 |
| Sheep | 7 | 13 | 6(7) | 4 | 16-18 |
| Goat | 7 | 13 | 6(7) | 5 | 16-18 |
| Swine | 7 | 14-15 | 6-7 | 4 | 20-23 |
| Camelid | 7 | 12 | 7 | 4 | 13-15 |
| Bird | 8-25 | 7 - four fuse to form notarium | Synsacrum - last 1-2 thoracic, plus the lumbar, sacral and first caudal vertebrae | 5-6 free vertebrae, then Pygostyle | |
In transverse section the spinal cord is tubular in shape, with a small central canal filled with CSF. It is divided anatomically into peripheral white matter and central grey matter. The butterfly-shaped grey matter is where somata are located; this is divided functionally into two or three main regions called horns. The horns form continuous columns of nerve cells that extend along the spinal cord. The white matter comprises tracts travelling cranially (ascending/sensory), caudally (descending/motor) and between segments (intersegmental). It is divided into anatomical regions called funiculi (Fig. 1.5). The spinal cord is covered in more detail in Chapter 4.
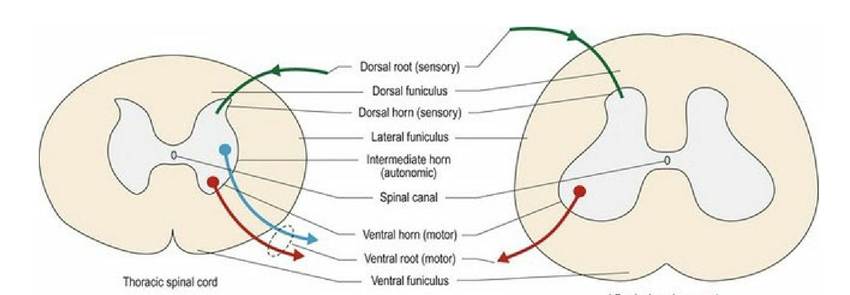
Fig. 1.5 Thoracic and lumbar spinal cords in transverse section depicting the basic arrangement of white and grey matter.
The basic anatomy and associated function of the different components of the spinal cord are outlined in Table 1.4.
Table 1.4 Functions of the different regions of grey and white matter in the spinal cord
| Greymaner | Function | White matter | Function |
| Dorsal hor∩ | Sensory | Dorsal funiculus | Ascending sensory (proprioceptive, tactile and nociceptive) Bidirectional, Intraspinal connections |
| Intermediate horn (thoracolumbar and sacral segments only) | Autonomic | Lateral funiculus | Ascending sensory (proprioceptive, tactile, nociception, thermal) Descending motor facilitating flexor muscle activity Bidirectional, Intraspinal connections |
| Ventral horn | Motor | Ventral funiculus | Descending motor facilitating extensor muscle activity Some ascending sensory (nociceptive) Bidirectional, Intraspinal connections |
The sulcus limitans is a small transverse fissure located laterally in the walls of the central canal of the spinal cord; it extends rostrally into the brainstem. This fissure arose during embryological development. It demarcates the dorsal, sensory component (alar plate) from the ventral, motor component (basal plate) (see Chapter 2).
Brain - forebrain, brainstem, cerebellum
The brain is housed in the protective neurocranium comprising frontal, parietal, occipital, temporal, ethmoid and sphenoid bones.
It can be divided anatomically and functionally. Anatomically there are three major divisions called the forebrain or prosencephalon, the midbrain or mesencephalon, and the hindbrain or rhombencephalon. These anatomical divisions may be subdivided further.
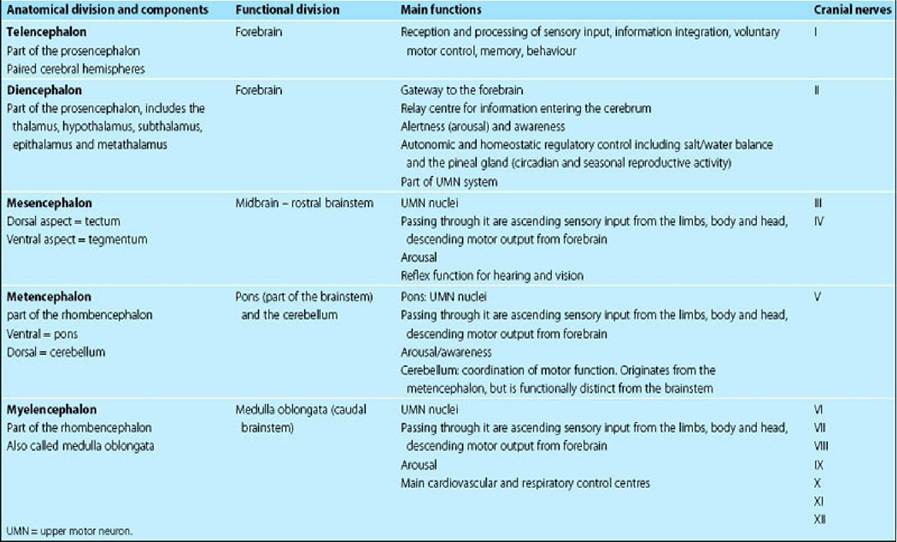
The three major functional divisions of the brain are the forebrain, brainstem and cerebellum. For functional reasons, we define the brainstem to include the midbrain, pons and medulla oblongata; however, some authors include the thalamus in the brainstem (Table 1.5).
Table 1.5 Regions of the brain, their general function and associated cranial nerves UMN = upper motor neuron
Anatomically, on the dorsal and lateral aspects, the brain is dominated by two large, ovoid cerebral hemispheres that are elongated in the longitudinal direction. Caudal to the hemispheres, on the midline and extending for a variable distance laterally, (species specific) is the cerebellum. The longitudinal fissure separates the hemispheres from each other, whilst the transverse fissure separates the hemispheres from the cerebellum (see Fig. A1).
Ventrally, the rostral aspect of domestic animals is dominated by the paired olfactory bulbs (Fig. 1.6A). These are small in humans (Fig. 10.3) and miniscule, or totally absent, in many cetaceans. Caudally, the olfactory bulbs are connected via the olfactory tract to the more laterally placed, piriform lobes. On the midline is the diencephalon with the optic chiasm, hypophysis (pituitary gland), and mammillary bodies (Fig 1.6B). Caudal to this is the midbrain with the crus cerebri (cerebral peduncles) conveying efferent information from the forebrain to the brainstem; the intercrural fossa is sited between the paired crura. Caudal to the midbrain is the pons of the ventral metencephalon, and then the medulla oblongata with the trapezoid body and the pyramids extending to the spinal cord junction.
Fig. 1.6A
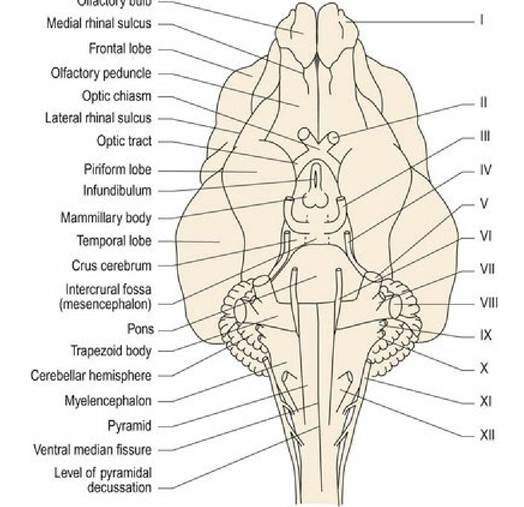
Canine brain, ventral aspect (see Fig. A3).
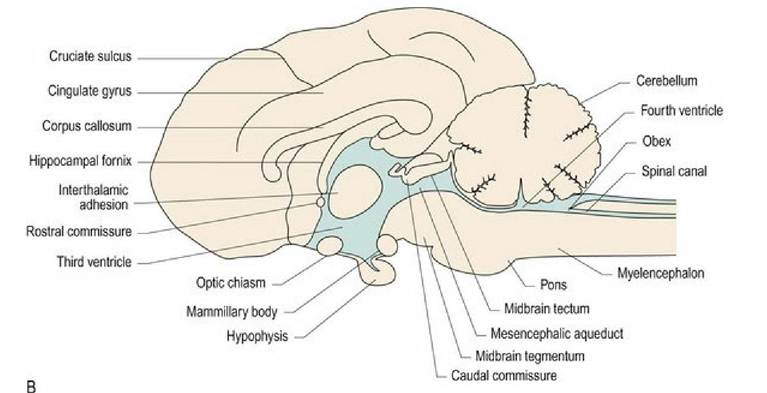
Fig. 1.6B Median section of canine brain (see also Figs. 3.11 and A4).
Brainstem
The brainstem forms a stalk connecting between the spinal cord and the cerebral hemispheres. The brainstem has a similar cross-sectional arrangement to that of the tubular spinal cord except that in the rostral medulla oblongata, the dorsal aspect of the tube has been opened up and the roof plate expanded (Fig. 1.7). The fluid- filled ventricular system is enlarged in this region, forming the fourth ventricle; this ventricle connects to the subarachnoid space via the lateral apertures (see Chapter 3). Rostral to the fourth ventricle is the mesencephalic aqueduct and caudally, the roof plate tapers and fuses on the midline at the site of the obex, reducing the ventricular system to the tube that continues caudally as the spinal canal.
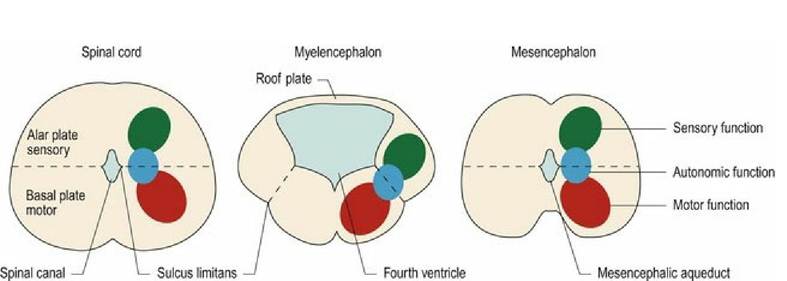
Fig- 1.7 Transverse sections of the spinal cord, medulla oblongata and midbrain and the function of the
nuclei in the different dorsoventral regions.
The white matter is superficial and the grey matter is located deeper within the brainstem. The sulcus limitans is more visible in parts of the brainstem than it is in the spinal cord. It still separates dorsal, sensory areas from the ventral, motor areas; however, in the fourth ventricle region it is displaced to a ventrolateral position. Autonomic (parasympathetic) grey matter is located at the level of the sulcus limitans at the intermediate position.
In the spinal cord, the grey matter forms continuous longitudinal columns, such that the sensory dorsal horn observed in cross-section is continuous along the length of the spinal cord. The same applies to the motor ventral horn. The autonomic nerve cells of the intermediate horn are present in the thoracolumbar area (sympathetic function) and the sacral area (parasympathetic function). The grey matter of the brainstem is similar to that of the cord in that it is longitudinally arranged, but the columns have become fragmented to form clusters of cell bodies with the same function. These clusters are called nuclei and many nuclei are associated with the cranial nerves (Fig. 10.2).
Nuclei are collections of nerve cell bodies, with similar functions, located within the CNS (Fig. 1.7). Ganglia are collections of nerve cell bodies, with similar functions, located outwith the CNS. Thus, it is inaccurate to refer to the ‘basal ganglia’ of the forebrain. The correct term is basal nuclei.
The dorsal aspect of the midbrain is associated with four bulges that together form the paired rostral and caudal colliculi (colliculus - L = hill). The two pairs of colliculi form the corpora quadrigemina (L = four bodies). The caudal colliculi are more widely separated from each other than are the rostral colliculi.
The reticular formation (rete - L = net) is a diffuse network of neurons extending from the medulla oblongata to the thalamus. It has a variety of functions including arousal/awareness, UMN function, cardiac and respiratory control centres.
Cerebellum
Prenatally, the cerebellum develops as a dorsal outgrowth of the metencephalon and adopts a complex structure. It has a highly convoluted surface with sulci (grooves) and folia (ridges). Grossly, it has a longitudinal, median ridge, the vermis, and two lateral hemispheres. The vermis is divided into approximately ten lobules (Fig. 7.1) that are associated with coordinating different aspects of motor function. For example, the caudoventral lobule, the nodulus and its adjacent hemispheres, the flocculus, together form the flocculonodular lobe. This lobe is also called the vestibulocerebellum as it connects to the vestibular nuclei of the brainstem. It is responsible for coordinating motor function to maintain the animal’s balance (see Chapter 7).
Forebrain
The forebrain comprises the cerebral hemispheres (cerebrum/telencephalon) and the thalamus (diencephalon). The thalamus acts like a gateway between the brainstem and the cerebrum such that all incoming information (except olfaction) approaches the cerebrum via the thalamus of the diencephalon. Thus the thalamus acts like a post office, receiving incoming mail from the brainstem, sorting it and sending it on to specific addresses within the cerebrum. It also receives local mail from the cerebrum and posts it back to other telencephalic areas. The diencephalon has five major subdivisions each associated with different functions, as outlined in Table 1.6.
Table 1.6 Regions of the diencephalon and their major functions
| Region of diencephalon | Anatomical location | Main functions |
| Thalamus | Midline: paired thalamic nuclei connected via the interthalamic adhesion, which passes through centre of third ventricle | Relay centre for incoming information, except olfaction Integration between cerebral areas Cerebral arousal |
| Hypothalamus | Ventral to the thalamus, forming the walls and floor of third ventricle Includes rostral (supraoptic) intermediate (tuberal) and caudal (mammillary) areas | Brain centre regulating autonomic motor activity |
| Subthalamus | Caudolateral to hypothalamus; ventral to the thalamus | Part of UMN motor control system |
| Metathalamus | Caudodorsal to thalamus; between the thalamus and the midbrain Includes the geniculate bodies | Relay centre for visual or auditory stimuli |
| Epithalamus | Dorso-medial to thalamus Includes the pineal gland and habenular nuclei. The latter has connections with the limbic system and olfactory regions | Circadian rhythms Autonomic responses to olfactory and emotional stimuli |
The cerebrum comprises two hemispheres (Figs. 1.8, A1). The surface of the hemispheres may be smooth (lissencephalic) or convoluted (gyrencephalic), forming gyri (ridges) and sulci (troughs) (Figs. 1.9, A2). The convolutions permit more cortical surface area to be contained in the same volume of cranial vault. The convoluted cortex is usual in most species of veterinary interest; however, rodents, lagomorphs, birds and most marsupials are naturally lissencephalic (Fig. 1.8). Developing carnivores and primates are lissencephalic until relatively late in gestation.
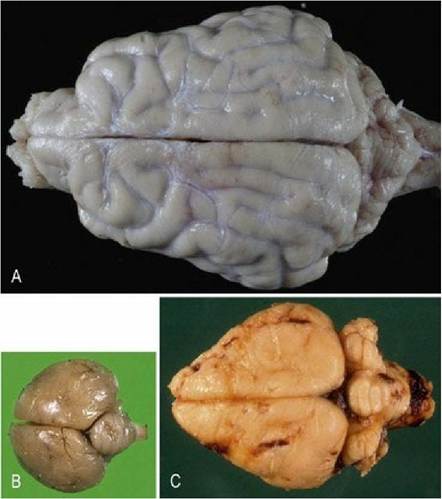
Fig. 1.8 Comparative anatomy of dog (A), bird (B) and rabbit brains (C) (dorsal aspects). Note the convoluted, gyrencephalic hemispheres in the dog brain, but lissencephalic hemispheres of the bird and rabbit brains. (Brains not to scale.)
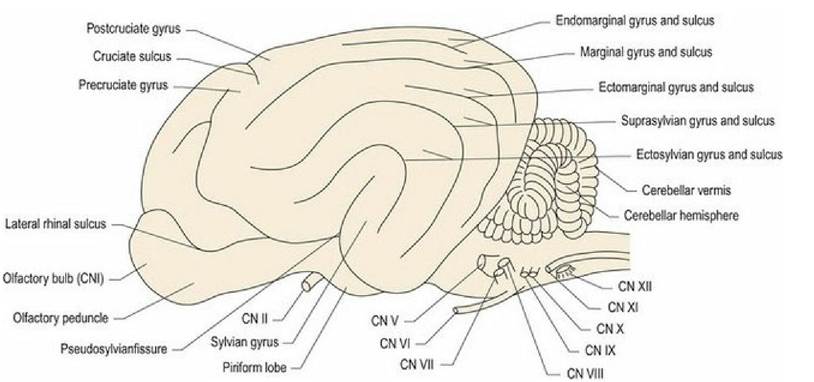
Fig. 1.9 Canine brain from the lateral aspect. Roman numerals refer to the cranial nerves. Note: the pattern of gyri and sulci is species specific.
Most of the grey matter is located superficially forming the cerebral cortex (Fig. 1.12). Grey matter is also located deep within the hemispheres in the hippocampus, the basal nuclei and septal nuclei. The hemispheres are divided into lobes named mainly for the overlying bone (Figs. 1.10 and 1.11 and Table 1.7). These lobes are loosely associated with different functions.
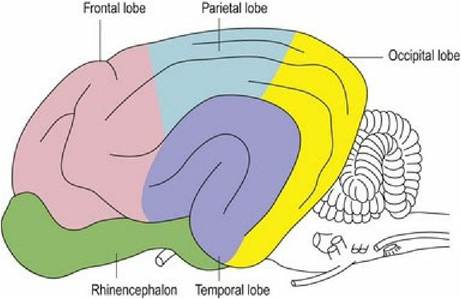
Fig. 1.10 Mammalian brain, lateral aspect, depicting lobes of brain.
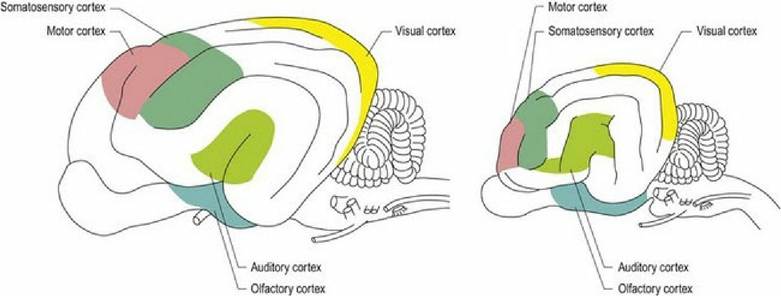
Fig. 1.11 Lateral aspect of the brains of the dog (left) and cat (right) depicting the main functional areas
of the brain.
Table 1.7 Lobes of the cerebrum and their major functions. Cognition is the ability to integrate a sensory input with other information such as memory. Thus an object that has been sensed can be recognised
| Region of cerebral hemisphere | Anatomical location | Main functions |
| Frontal lobe | Deep to the frontal bone, forming rostral third of the hemisphere | Motor cortex, association areas involved in planning actions and movement |
| Parietal lobe | Deep to the parietal bone. Dorsal midline strip caudal to frontal lobe, extending one-third of the way down lateral aspect | Somatosensory (somesthetic) cortex and association areas Cognitive association areas involved in perceiving sensory input |
| Temporal lobe | Deep to the temporal bone Ventrolateral mid portion of cerebral hemispheres | Audition, limbic system, learning and memory |
| Occipital lobe | Deep to the occipital bone, caudal third of hemispheres | Vision Cognitive association areas |
| Rhinencephalon | Ventro-lateral aspect of each hemisphere, ventral to lateral rhinal sulcus, dorsal to the sphenoid bones and caudal to ethmoid bone. Includes the olfactory bulbs, piriform lobe and hippocampus | Olfaction, limbic system, learning and memory |
In vertebrate animals, brain weight is closely correlated with body weight, thus an adult sperm whale has a brain that weighs five times more than that of a human. However, the white matter increases as a cubic function while the surface area and grey matter content only increase as a square function: thus brains from larger animals have a higher white matter to grey matter ratio. In an evolutionary sense, the cortex is divided into primitive, old and new regions. These are the paleopallium (paleo, derived from the Greek for ancient; pallium - L = cloak), archipallium (archi - Gk = primitive) and the neopallium (neo - Gk = new). The paleopallium is ventral to the lateral rhinal sulcus and is largely associated with olfaction. It is relatively large in domestic animals compared with primates. The archipallium, which includes the hippocampus, is a region of the brain that originally functioned to correlate olfactory input with other sensory information, but evolved to acquire other functions such as behaviour (see Chapter 11). It is medially located. The neopallium is the largest part of the cerebrum, separated from the archipallium by the splenial sulcus medially and from the paleopallium by the lateral rhinal sulcus, ventrally (see Figs. A2, A4). It includes the primary receiving areas for vision, audition and sensory inputs, as well as integration areas.
White matter tracts form connections within the forebrain (association and commissural fibres) and to other parts of the CNS (projection fibres) (Fig. 1.12). Association fibres connect within a hemisphere. Short association fibres connect within a lobe between adjacent gyri, while long association fibres connect between lobes within the same hemisphere. Commissural fibres connect across the midline between hemispheres. The main commissural fibres are bundled together as the corpus callosum. This forms an elongated, transverse band of white matter at the base of the longitudinal fissure. Projection fibres connect between the hemispheres and the brainstem (Fig. 1.13). Corticopetal and corticofugal projection fibres enter and leave the cerebrum, respectively; they do so via the internal capsule.
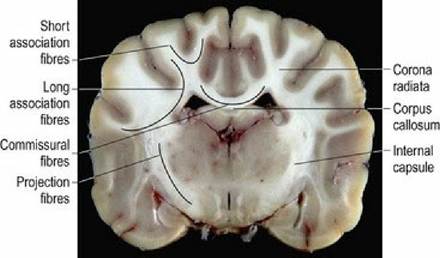
Fig. 1.12 Dog brain, transverse section, at the level of the thalamus, illustrating white matter connections.
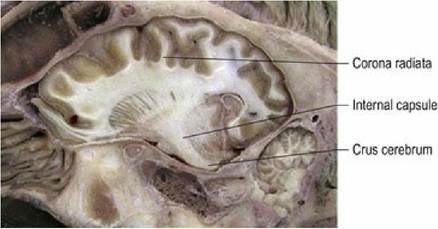
Fig. 1.13 Sheep brain, paramedian section, depicting white matter connections of the cerebrum to the
brainstem via the internal capsule
(courtesy of the Veterinary Virtual Museum, IVABS, Massey University).