Metabolism
The terms anabolic and catabolic are used to describe the overall metabolic state or status of an animal. Anabolism refers to a constructive process (e.g., synthesis of proteins from amino acids), while catabolism refers to a destructive process (e.g., degradation of proteins into individual amino acids).
While anabolic and catabolic refer to the overall metabolic status, both processes are usually ongoing at the same time in an animal’s body. For example, even in a young, rapidly growing animal that is digesting a meal, absorbing nutrients, and synthesizing body proteins, some body proteins are degrading at the same time. However, in this case, the rate of protein synthesis is greater than the rate of protein degradation, so the animal is in an anabolic state.several hormones contribute to the regulation of the balance between anabolic and catabolic processes, and the study of the hormones that regulate metabolism is termed metabolic endocrinology. A common feature of such hormones is that blood glucose concentrations participate in regulation of their secretion. This indicates that the maintenance of a minimal and constant source of glucose for energy is a key factor in the overall endocrine control of metabolism.
To study the endocrine control of anabolism and catabolism, metabolic endocrinologists often contrast the period shortly following a meal, during which nutrients are being absorbed from the gastrointestinal tract (absorptive state), with a period during which there is no net absorption (postabsorptive state). During the absorptive state, blood levels of glucose, amino acids, and triglycerides (as part of chylomicrons) increase. The overall goals of the metabolic processes during this period appear to be to increase the use of these nutrients by cells of the body or store them so that they can be used later. Metabolic periods of ruminants differ from those of other animals, because nutrients are constantly being absorbed from the forestomach and passing from the forestomach down the remainder of their gastrointestinal tract.
This chapter discusses some specifics of ruminant metabolism after a more general overview.Absorptive State: Anabolism
Figure 22-1 summarizes the overall fate of the major nutrients absorbed during the digestion of a meal, and these are described in more detail in the following paragraphs.
Glucose is the predominant product of carbohydrate digestion in most animals, and following a typical meal, blood glucose levels may rise to 150% of fasting levels. The increase in blood glucose is a major stimulus for the release of insulin from the pancreas, but increases in plasma amino acids during the digestion of a high-protein meal can also stimulate insulin release. insulin affects carbohydrate, amino acid (protein), and lipid metabolism during the absorptive period, and it is considered the primary endocrine regulator of metabolism during anabolism.
insulin stimulates the uptake of glucose by skeletal muscle cells, where it can be used for energy or stored as glycogen (essentially a polymer of glucose molecules). The liver also stores glucose as glycogen during the absorptive
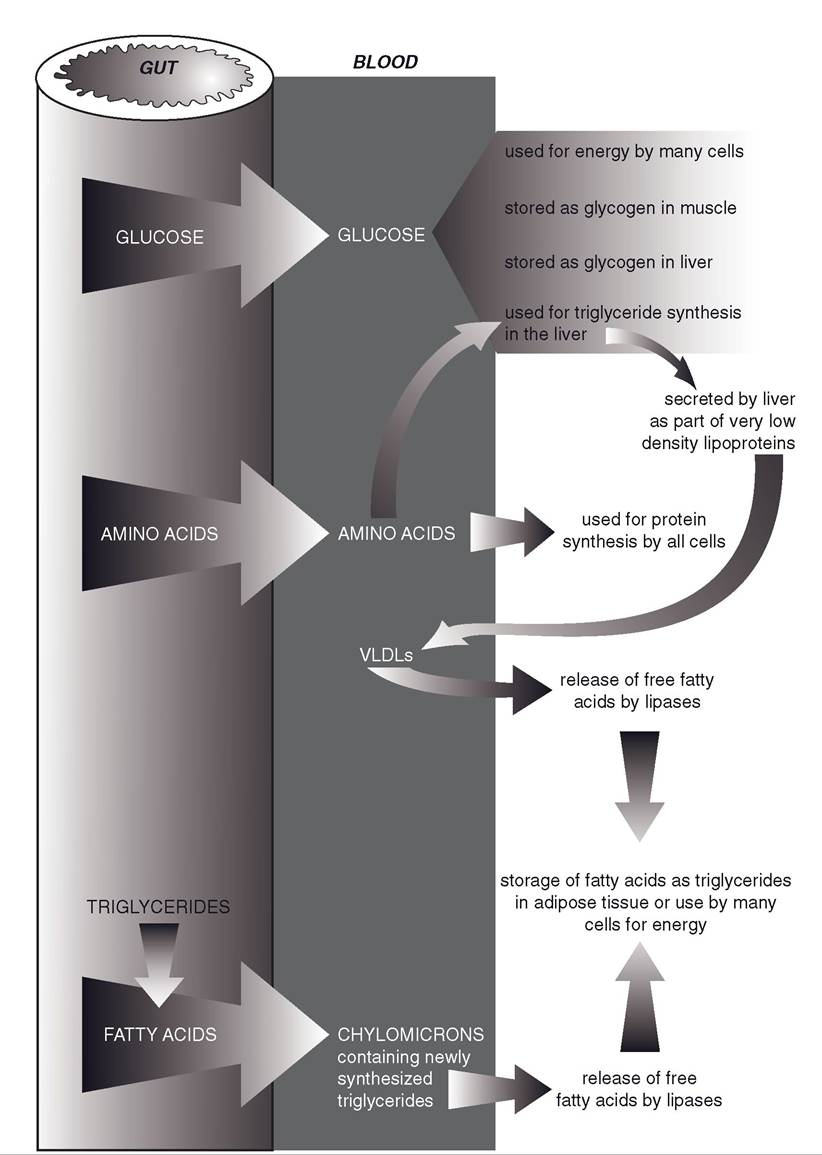
Figure 22-1. Metabolic fate of glucose, amino acids, and triglycerides (in chylomicrons) being absorbed from the gastrointestinal tract during the digestion of a meal.
period, and this is also stimulated by insulin. Primarily because the mass of skeletal muscle is greater than the liver, much more glycogen (75% of the total) is formed and stored in skeletal muscle. Blood glucose is also available for use by all cells of the body for energy during this period, but no other organ is capable of significant glycogen storage.
Absorbed amino acids are immediately available to all body cells for protein synthesis. Because all of the amino acids necessary for synthesis of a given protein must be available at the time of synthesis, it is imperative that animals have a balanced diet that contains all of the essential amino acids.
Protein synthesis in many organs, including liver and skeletal muscle, is stimulated by insulin, so the increase in insulin following a meal also promotes protein synthesis during this period. However, this stimulatory effect on protein synthesis is minor compared to the effects of insulin on glucose metabolism (e.g., the rate of plasma protein production by the liver increases by only a small percentage after a meal).The amounts of amino acids absorbed after a typical meal are more than can be efficiently used by the body for protein synthesis. However, no metabolic pathways permit the various amino acids to be stored for later use the way glucose is stored as glycogen. Many of the excess amino acids are taken up by hepatocytes and enter metabolic pathways that result in triglyceride (lipid) formation. These pathways remove nitrogen-containing amino groups from the amino acids (deamination). Most of the resulting lipids are secreted by hepatocytes into the blood as part of lipoproteins (discussed later).
Deamination of amino acids is also part of a different metabolic pathway by which liver cells use amino acids to produce glucose. However, the hormone glucagon must be available to stimulate this pathway, and glucagon release from the pancreas is reduced by increases in blood glucose. Thus, during the period that blood glucose is elevated following a meal, the use of amino acids to produce glucose is suppressed. Gluconeogenesis is the term for the collective metabolic processes by which liver cells produce glucose from non-carbohydrate substrates, such as amino acids and short-chain fatty acids. The liver and kidneys are the only organs that are capable of any significant gluconeogenesis, and the kidneys do so only in states of chronic acidosis.
During the absorptive state, the liver uses both excess glucose and amino acids as substrates for triglyceride (lipid) synthesis, and insulin stimulates these pathways. some of the newly synthesized triglycerides are stored in the liver, but most are released into the blood in complex particles known as very low density lipoproteins (VLDLs).
Lipoproteins are particles that contain lipids, cholesterol, and proteins in various ratios. VLDLs are so named because their lipid content is high relative to their protein content. Because lipids are less dense than protein, the density of VLDL particles is quite low.Recall that chylomicrons are also circulating lipoproteins, but the triglycerides in these lipoproteins were absorbed from the intestinal tract. As chylomicrons and VLDLs circulate throughout the body, they encounter lipoprotein lipase, an enzyme bound to endothelial cells that acts on their triglycerides to release free fatty acids. When the triglycerides are released within adipose tissue, the free fatty acids are available to adipose cells for the resynthesis and storage of lipids as triglycerides. in other organs, such as skeletal muscle, cells use the free fatty acids for energy. The synthesis and storage of triglycerides in adipose tissue is stimulated by insulin, which is typically elevated during the absorptive period.
After losing triglycerides by the action of lipoprotein lipase, some VLDLs undergo changes in the circulation and become a different type of lipoprotein, low-density lipoprotein (LDL). LDLs contain a great deal of cholesterol, and cells throughout the body receive cholesterol from the blood by the endocytosis of LDLs. Cholesterol is a necessary component of cell membranes, and all cells need some cholesterol. However, abnormal increases in LDL levels are associated with an increased risk of cardiovascular disease in humans. Recall that the original
VLDLs were produced in the liver, so much of the cholesterol in the blood is produced by the liver.
Postabsorptive State: Catabolism
After a meal has been digested and absorbed, blood glucose concentration gradually decreases
as glucose is used for energy throughout the body (Fig 22-2). This drop in blood glucose is the primary event bringing about the changes in endocrine secretions that orchestrate the metabolic changes during the postabsorptive state.
Two major endocrine changes are a gradual drop in insulin secretion and a rise in the release of glucagon. Recall that increases in blood glucose stimulate insulin release from β-cells, whereas decreases in blood glucose stimulate glucagon release from α-cells in pancreatic islets.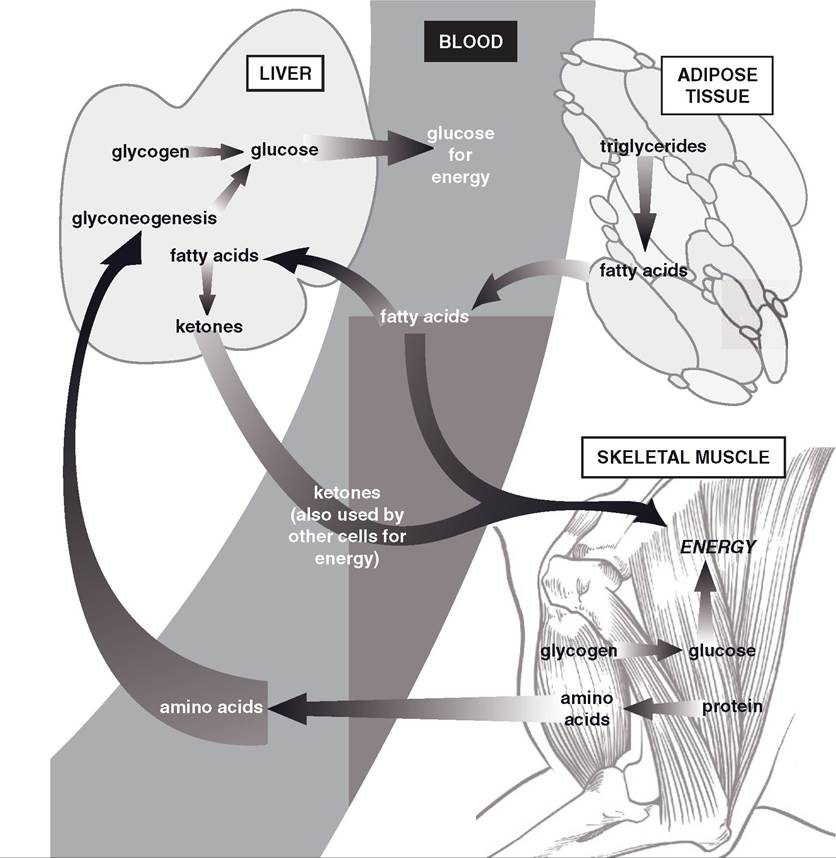
Figure 22-2. Summary of metabolic organs and mechanisms that maintain blood glucose and provide energy for cells when nutrients are not being absorbed from the gastrointestinal tract.
During the absorptive period, when glucose and amino acids were being absorbed into the blood from the intestinal tract, insulin stimulated the synthesis of glycogen for glucose storage, proteins, and lipids (from any excess glucose and amino acids). As insulin levels decrease, its stimulatory effect on these synthetic (anabolic) processes is lost, and this is a major factor in changing the overall metabolic balance from anabolism to catabolism.
Glucagon stimulates the breakdown of glycogen (glycogenolysis) in the liver to provide glucose that the liver can release into the blood. Glycogenolysis is the initial process by which the liver derives glucose to add to the blood, but later the liver also releases glucose formed by gluconeogenesis, which is also stimulated by glucagon. The amino acids used for gluconeogenesis are derived from the catabolism of body protein.
The maintenance of a minimal or fasting blood glucose level during this period has primary importance to neuronal function. Neurons do not have the metabolic processes to permit them to use fatty acids for energy, so they need a ready supply of glucose for cell energy. During fasting, the catabolism of adipose tissue increases the supply of fatty acids, which are used by cells other than neurons for energy. The increased use of fatty acids by other cells reduces the overall need for glucose and conserves it for use by neurons.
This conservation of glucose is termed glucose sparing. The liver also metabolizes the circulating fatty acids to produce ketones, another cellular energy substrate. The ketones produced by the liver include acetone, acetoacetate, and β-hydroxybutyrate.Two other hormones, growth hormone and glucocorticoids from the adrenal cortex, also contribute to the maintenance of blood glucose and other sources of energy during anabolic periods. Glucocorticoids do not increase in the circulation during a short fast, but a deficit of glucocorticoids reduces the rate of liver gluconeogenesis and mobilization of fatty acids from adipose tissues. The effect of glucocorticoids on these processes during fasting is a permissive effect. Decreases in blood glucose stimulate the release of growth hormone, which increases the mobilization of fatty acids from adipose (lipolysis). Some tissues (e.g., skeletal muscle) can use the fatty acids for energy (glucose sparing), and the liver can further increase its production of ketones.
Energy Needs During Exercise
The increase in skeletal muscle metabolism during exercise can rapidly deplete the glycogen stores within skeletal muscle cells. in humans these stores are capable of providing energy only for an estimated 2 to 3 minutes of very intense exercise. To sustain exercise, other energy sources must be rapidly mobilized and delivered to working skeletal muscle. Circulating levels of epinephrine and norepinephrine increase during exercise, and these catecholamines have several actions that mobilize energy stores, including increased glycogenolysis in the liver and nonworking skeletal muscle and lipolysis in adipose tissue. insulin levels are reduced and glucagon levels are increased during exercise, which promotes liver gluconeogenesis and more lipolysis in adipose tissue. The decrease in insulin is not detrimental to working skeletal muscle, because glucose uptake by working muscle is less insulin dependent.
Anaerobic metabolism, by working skeletal muscles, raises the rate of lactic acid production. The lactate ion can diffuse into the blood from the skeletal muscle, and plasma levels of lactate increase during strenuous and prolonged exercise. The liver can use blood lactate for gluconeogenesis, and glucose can then be returned to the blood to maintain blood glucose levels.
Blood Glucose in Ruminants
Normal ranges for blood glucose levels in mature ruminants are lower than in other animals, even other herbivores (cattle, 45 to 80 mg/dL; dog, 70 to 110mg/dL; and horse, 60 to 110 mg/dL). The lower normal range for mature ruminants is associated with the relatively small amount of glucose-yielding carbohydrate digestion in their small intestine. Most of the carbohydrates that they consume undergo fermentative digestion in the forestomach and result in the production of short-chain volatile fatty acids (VFAs), which are absorbed directly from the forestomach.
Without glucose readily available via absorption from the gastrointestinal tract, ruminants must have a continuous and a relatively high rate of gluconeogenesis in the liver to maintain the blood glucose level. Glucagon appears to be an important endocrine stimulant to maintain this rate of gluconeogenesis. Rising levels of amino acids and propionic acid (a VFA produced in the rumen) can stimulate glucagon release, so presumably the continual absorption of these from the ruminant gastrointestinal tract can maintain glucagon secretion. Propionic acid is the one of the three major VFAs produced in the rumen that can be used by the liver for gluconeogenesis.
Ketosis
Ketosis is a metabolic state characterized by an increase in blood ketones, a reduction in urine and blood pH, and ketones in the urine. The increase in the acidic ketones in the blood and urine are responsible for the changes in pH. Ketosis may occur when fatty acid mobilization from adipose tissue is elevated and glucose is deficient. The deficiency in glucose stimulates the release of glucagon and inhibits insulin release, and the increased ratio of glucagon to insulin promotes the formation of ketones by the liver from readily available fatty acids.
Ketosis may develop in dairy cattle at the peak of lactation, when the need for glucose to synthesize lactose (milk sugar) is maximal. The rapid use of glucose by the mammary glands reduces blood glucose and brings about these changes in glucagon and insulin. Ketosis may also develop as a result of type i diabetes mellitus, in which the primary problem is a deficiency of insulin. in this case, the dominant effects of glucagon on fatty acid mobilization and ketone synthesis are primarily responsible for the development of the ketosis.