Neural Control of Skeletal Muscle
As described in Chapter 8, each contraction of skeletal muscle requires stimulation of the muscle by a somatic motor neuron. somatic motor neurons have their cell bodies within the CNS, and their axons extend from the CNS to contact skeletal muscle cells at neuromuscular junctions.
Each time an action potential passes down the axon of a single motor neuron, all skeletal muscle cells that are innervated by that motor neuron are stimulated to contract. A single motor neuron can stimulate more than one skeletal muscle cell, but a single skeletal muscle cell receives input from only one motor neuron. Thus, all contractions of any single skeletal muscle cell must be initiated by action potentials arriving via the same motor neuron. A clinical classification applied to the motor neuron that extends from the CNS to skeletal muscle fibers is lower motor neuron (LMN). (More specifically, this is the somatic LMN, but somatic is often omitted in common usage.) Any contraction of skeletal muscle depends on an intact and properly functioning LMN, and the classic sign of a dysfunctional LMN is flaccid paralysis (a total lack of movement and muscle tone of the affected muscle).Typically, a single neuron receives inputs from multiple other neurons because of convergence in neural networks. Therefore, a single LMN is subject to regulation by multiple inputs from other neurons within the CNS. Upper motor neuron (UMN) is the general term applied to a neuron within the CNS that acts to regulate the activity of a LMN. UMNs are responsible for initiating voluntary movement and maintaining relatively stable body posture and position relative to gravity so that voluntary movements are normal. if the LMNs to a muscle are intact but some of the UMNs that normally regulate those LMNs are dysfunctional, both reflex and voluntary activity of the muscle are possible, but the character of both reflexes and voluntary activity may be abnormal.
Reflexes Involving Skeletal Muscle Contraction
A refl ex action is an automatic, or unconscious, response to an appropriate stimulus. in the case of reflexes involving skeletal muscle, the response is skeletal muscle contraction. The simplest reflex requires two neurons. The first is an afferent neuron, which detects the appropriate stimulus and transmits that information via action potentials to the CNS. Within the CNS the afferent neuron synapses with and stimulates action potentials in an efferent neuron. The efferent neuron exits the CNS and stimulates skeletal muscle contraction. The entire neural circuit from stimulus detection to response is a refl ex arc, and in the simplest case, in which only one synapse is involved, the reflex arc is a monosynaptic reflex (Fig. 10-5). Very few reflexes are monosynaptic; usually one or more interneurons are interposed between the afferent and efferent neurons.
Reflexes can be classified according to the segments or parts of the CNS that are required
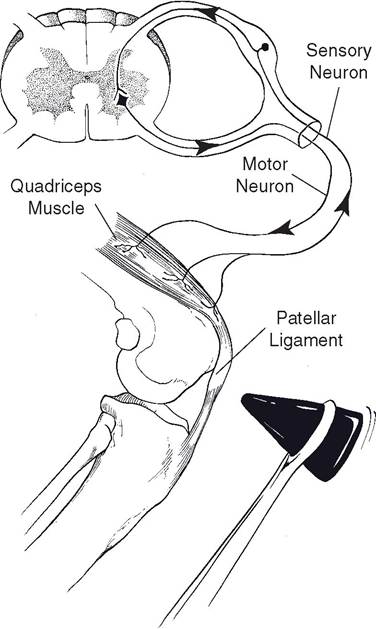
Figure 10-5. Patellar tendon reflex arc is a monosynaptic spinal reflex.
to complete the reflex arc or circuit. Among the simplest of reflex arcs are those that can be completed within segments of the spinal cord (spinal reflexes). A typical spinal reflex is the patellar tendon reflex, or knee jerk reflex (Fig. 10-5). The knee jerk is elicited by tapping the patellar ligament (tendon), which causes the knee or stifle to extend. The sudden stretching of the muscle stimulates muscle spindles (see Chapter 11) related to the quadriceps muscle. Action potentials are transmitted along axons of primary afferent neurons and into the spinal cord by way of the dorsal root. A synapse occurs between the afferent neuron and appropriate motor neurons in the ventral gray horn of the spinal cord. Action potentials then are transmitted via the efferents to the muscle fibers of the m.
quadriceps femoris of the thigh (in this case the muscle in which the stimulus originated), causing it to contract. This type of stretch reflex is also known as a postural reflex because it aids in maintaining a standing position. sudden flexing of the stifle stretches the quadriceps muscle, which initiates the reflex that causes the stifle to support the weight of the animal. The same reflex occurs in humans when the area just below the knee is struck.The fact that the patellar tendon reflex arc can be completed within a limited region of the spinal cord does not mean that other segments of the CNs cannot affect the characteristics of the reflex response. Through the actions of either inhibitory or excitatory uMNs that affect the activity of the LMNs participating in the reflex arc, the strength or character of the reflex response can be changed. As described earlier, the patellar tendon reflex is an example of a postural reflex that functions to maintain a normal posture and body position. By affecting the character of the reflex, uMNs can also assist in the maintenance of posture and body position. More information about the regulation of uMNs is given later in this chapter.
Reflex center is a general term used to describe the area within the CNs where afferent information is integrated to produce the efferent activity or reflex response. in the case of simple spinal reflexes, the reflex center is within the spinal cord (i.e., the synapses are between the afferent and efferent neurons). Reflexes can be quite complex and may require integration by reflex centers within the brain or brainstem. For example, vomiting is a reflex that can be stimulated by a variety of diverse stimuli and that requires a series of highly coordinated contractions of skeletal muscles in the diaphragm, esophagus, and pharynx. These actions are coordinated by a reflex center in the brainstem. This center receives afferent information from quite diverse areas (such as the stomach lining and the vestibular apparatus) and stimulates efferent neurons to all participating skeletal muscles.
This center can be stimulated pharmacologically by the administration of any of a variety of emetic drugs.in the dog, spinal refl exes that are used in various types of clinical examinations include the withdrawal refl ex, in which the limb is withdrawn when the footpad or toe is pinched; patellar tendon reflex, in which extension of the stifle results from tapping the patellar ligament; and extensor postural thrust refl ex, in which the limb extends when the foot is pushed toward the body, as would occur in normal body support.
Attitudinal and postural reactions include the tonic neck refl exes, in which passively extending the neck (raising the head) increases the tone of extensor muscles of the forelimbs and decreases the tone of extensor muscles of the hind limbs. This may be a partial explanation for the value of raising a horse’s head to prevent kicking. Conversely, when a horse gets its head down, it is much more likely to kick or buck. The extensor postural thrust enables the limbs to support the weight of the body without conscious thought on the part of the animal.
Reflexes associated with the cranial nerves include the corneal refl ex, in which the eyelids close in response to stimulation of the cornea of the eye, and the palpebral refl ex, in which gentle palpation or tapping of the eyelids causes closure. General anesthesia depresses reflex responses, and these last two reflexes are often used to evaluate the depth of anesthesia.
Voluntary Movement and Its Regulation
Animals cannot be asked about their intentions or thoughts while moving about. Therefore, much of what is known or assumed about voluntary movement in animals is based on information from humans; that is, humans are the experimental animals. Much of the information from humans is based on studies of individuals with naturally occurring diseases or acquired neural deficits due to trauma. Studies on animals with similar lesions or deficits suggest that the underlying mechanisms are the same in humans and animals.
Using a variety of techniques and experiments, a large area within the cerebral cortex has been shown to be directly linked to motor activity (i.e., the motor cortex). Within this general area, more specific areas can be shown to control various groups of skeletal muscles. While this does not mean that neurons from this area make direct synaptic connections with skeletal muscle cells, a functional association between these areas and specific skeletal muscles can be demonstrated by eliciting contractions of specific muscles by stimulation of discrete areas within the motor cortex. Groups of skeletal muscles used for fine motor activity (e.g. fingers in humans) have a greater area of representation in the motor cortex than groups of muscles used for less-fine motor activity (e. g., bending over at the waist). The sizes of the areas in the motor cortex that represent different regions of the body are quite variable among different animals and between humans and domestic animals. For example, a horse would have a very small area representing the digits, whereas this would be large in humans. This difference is related to the different needs for fine motor control of the digits between horses and humans.
The precise linkage between the desire for and initiation of a specific voluntary movement (e.g., reaching for, grasping, and lifting a cup of coffee) and how the necessary muscles are appropriately stimulated to bring about that movement is not completely understood. What is understood about normal voluntary movement, and is more relevant to animals, is that it requires a great deal of afferent information before and during the movement, and this information is used for the coordination of voluntary movements. This afferent information is primarily derived from proprioceptors (specialized sensory receptors designed to provide information about body position discussed in Chapter 11) in muscles, tendons, and joints throughout the body. This information is vital for coordinated movements.
For example, to reach for a cup of coffee and grasp it, the position of arms, hands, and fingers must first be known. When the cup is lifted, continuous adjustments must be made in muscle contraction so that the movement is smooth regardless of the weight of the cup or the amount of coffee in it. Much of the afferent information on body position is received and used by the brain without any conscious awareness.The cerebellum receives much of the afferent information about body position and ongoing movements from proprioceptors. it also receives information about movements initiated in the cerebral cortex. After integrating this and other information, the cerebellum sends efferent information to multiple sites in the brain, including the motor cortex, to coordinate ongoing movements. The cerebellum does not initiate movements, but it is essential for normal coordination of voluntary movements.
it is clear that repetition of a pattern or sequence of movements increases the ease with which those movements can be repeated (i.e., training). This indicates that a type of learning can also occur within the neural circuits regulating voluntary movements. After such learning occurs, a series of practiced movements are virtually automatic and can be done without any conscious direction. Exactly how and where this learning occurs is not known, but the basal nuclei (see Chapter 9) appear to have special importance in coordinating and enabling the automatic performance of repeated movement patterns. Dopamine is an important neurotransmitter in certain basal nuclei, and Parkinson’s disease in humans is associated with the loss of dopaminergic neurons in this area of the brain. Patients with Parkinson’s disease exhibit a variety of abnormal movements, and complex movement patterns seem to be especially affected. Treatment with L-dihydroxyphenyalanine (L-dopa), a precursor of dopamine, may relieve the symptoms, but it does not prevent further neuronal loss and progression of the condition.
All UMNs have their effects on skeletal muscle activity and movement indirectly by ultimately affecting LMN neuron activity. Because the final common pathway for all movement (reflex or voluntary) is via the LMN and because the function of UMNs is to regulate LMN activity, abnormal movements may result from dysfunction of either LMNs or UMNs regulating the activity of a specific muscle. The abnormal movement may be primarily voluntary movements, reflex-based movements, or both. By close observation of abnormal movements and with a detailed knowledge of LMNs, UMNs, and the primary functions of the sites within the CNS from where UMNs originate, clinicians can determine the type and location of dysfunctional neurons causing the abnormal movements.