Physiology of Lactation
Composition of Milk
Milk contains all of the nutrients necessary for survival and initial growth of mammalian neonates. The nutrients in milk include sources of energy (lipids and carbohydrates), proteins to provide amino acids, vitamins, minerals (ash) for electrolytes, and water.
The relative amounts of these nutrients in milk vary among species (Table 29-1).Diet and the stage of lactation also affect the composition of milk. Diets high in non-fiber carbohydrate sources of energy are associated with increases in the percentage of lipids in the milk. Diets high in protein promote a slight increase in the percentage of protein in the milk, but this effect is much less than the effect of energy on milk lipid content. The amount of carbohydrates in milk (lactose, or milk sugar) does not routinely change with diet. The percentage of lipids and protein in milk is also highest early in lactation. in cattle the percentages are relatively high in the first few weeks after calving and then decrease over the next 3 to 4 months. Later in lactation, the concentrations of lipids and proteins again increase as total daily production (pounds of milk per day) decreases.
| Table 29-1. Typical Values for Constituents of Milk in Grams per Liter | |||||
| Species | Lipids | Lactose | Protein | Total Minerals (ash) | Calcium |
| Cow | 38 | 48 | 37 | 7.0 | 1.3 |
| Mare | 16 | 50 | 24 | 4.5 | 1.0 |
| Ewe | 70 | 40 | 60 | bgcolor=white>8.01.9 | |
| Sow | 80 | 46 | 58 | 8.5 | 2.0 |
| Doe | 40 | 45 | 35 | 7.8 | 1.2 |
Most of the lipids in milk are in the form of triglycerides, and these are the primary source of dietary energy in milk.
Triglycerides are composed of three fatty acids and glycerol. The fatty acids for the synthesis of milk triglycerides may be derived from the blood or synthesized within the mammary gland. Nonruminant mammary glands use blood glucose both for energy and as a source of carbon for the synthesis of the fatty acids. The glycerol is derived mostly from glucose catabolism in the process of glycolysis. The mammary glands of ruminants depend on blood acetate and β- hydroxybutyrate to provide carbon for fatty acid synthesis, with acetate being the primary source. The acetate and β-hydroxybutyrate in ruminants are produced as volatile fatty acids by fermentative metabolism by microorganisms in the rumen. These volatile fatty acids are absorbed into the blood and thereby become available for synthesis of milk fat in the mammary gland. Most milk triglycerides have fatty acids with chains 4 to 14 carbon atoms in length—short-chain fatty acids. Such shortchain fatty acids are not generally found in adipose tissue throughout the remainder of the body.Lactose (milk sugar), the principal carbohydrate in milk, is a disaccharide composed of the two monosaccharides glucose and galactose. Lactose is synthesized in mammary glands and is typically found only in mammary glands and milk. secretory cells in mammary glands use glucose from the blood to synthesize galactose and then combine the galactose with more glucose to produce lactose, so glucose is essential for lactose synthesis. The extraction of glucose from blood by an actively secreting mammary gland is quite effective, so the glucose concentration of venous blood leaving a mammary gland is relatively low.
Recall that in ruminants blood glucose is primarily derived from gluconeogenesis in the liver using propionic acid, a volatile fatty acid absorbed from the rumen, as a substrate. Thus, propionic acid produced by ruminal microorganisms and fermentative metabolism is the ultimate substrate for the production of lactose in ruminants.
Also, blood glucose is relatively low in ruminants compared to other mammals, in part because ruminants absorb very little glucose from the gastrointestinal tract. At peak lactation of a high-producing dairy cow, the mammary glands use most of the glucose produced by the liver for lactose production. If the need for glucose by the mammary glands cannot be met by gluconeogenesis, and blood glucose levels drop significantly, lactational ketosis develops. While blood glucose levels are low, metabolic acids (produced in the liver from fatty acids) accumulate in the blood to produce a metabolic acidosis.The major milk proteins are the caseins. Blood amino acids are the precursors for direct synthesis of the caseins by secretory cells within mammary glands. other milk proteins include α-lactalbumin and β-lactoglobulins, which are produced by cells of the mammary gland, and serum albumin and immunoglobulins produced by the liver and lymphocytes, respectively.
Rennin (also known as chymosin) is a proteolytic enzyme secreted by gastric epithelial cells of very young mammals. Rennin changes the character of ingested milk from a liquid to a semisolid curd; this process is termed curdling or coagulation. The change in character increases the time that milk is retained in the stomach, and this permits protein digestion to begin. Curdling results when rennin degrades one of the casein proteins responsible for increasing the solubility of micelles, casein protein aggregates in milk (discussed next). Without this specific casein, milk proteins precipitate with the calcium in milk to form curds.
Milk Secretion
The epithelial cells lining the alveoli of mammary glands are the cells primarily responsible for the secretion of milk. The appearance of these cells varies as they synthesize and release the lipids, proteins, and lactose of milk. After the cells actively secrete the constituents of milk and the lumen of the alveoli are filled with milk, the epithelial cells shrink and are described as a simple low-cuboidal epithelium (Fig.
29-5). At this stage, their secretory activity is relatively low. shortly after the stored milk is removed, the epithelial cells increase their secretory activity and begin to refill the alveoli. Early in the secretory phase the cells assume a more columnar appearance and then gradually reduce to cuboidal as milk fills the alveoli. small, apparently nonfunctioning alveoli can be found in dry mammary glands, and there is a relative increase in the amount of interstitial loose connective tissue (Fig. 29-4).Milk lipids are synthesized and packaged into secretory droplets, which are extruded from the luminal surface of the cell into the alveoli (Fig. 29-6). As they are released, a mem-
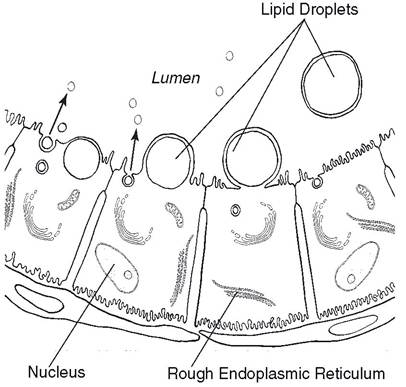
Figure 29-6. Secretion of milk lipids, milk proteins, and lactose by epithelial cells lining the alveoli of the mammary glands. Proteins and lactose are together in secretory vesicles that are released by exocytosis (arrows).
brane covering derived from the cell membrane of the epithelial cell encases the lipid droplets. The alveolar secretory cells also produce secretory vesicles that contain both milk proteins (caseins) and lactose (Fig. 29-6). As caseins are synthesized and packaged in these vesicles, they self-aggregate into particles termed micelles. The inclusion of one specific type of casein (κ-casein) in this aggregation increases the solubility of the micelle so that milk proteins remain in solution after their release from the cell. The lactose within the secretory vesicle generates an osmotic force to draw water into the vesicle from the cytosol of the cell. The secretory vesicles, each containing a mixture of micelles, lactose, and water, are transported to the apical surface of the cell and released into the alveoli by exocytosis (Fig. 29-6). Because of the various mechanisms by which lipids, proteins, and lactose are secreted from alveolar cells, milk is considered to be a combination of apocrine and merocrine secretions.
Lactogenesis
Lactogenesis is the establishment of milk secretion, and galactopoiesis is the continued production of milk by the mammary glands. Growth and development of the secretory epithelium and the ductile network of the mammary glands must precede lactogenesis. The initial extensive development of the mammary gland is usually associated with puberty (the beginning of sexual maturity) and the subsequent cyclic changes in the ovarian hormones, estrogen and progesterone. Estrogen particularly promotes the growth of the ductile system at each estrus, while progesterone, acting with estrogen, is required for growth and anatomic development of secretory alveoli. Normal secretion of growth hormone and glucocorticoids are also required for complete mammary gland development.
During pregnancy, prolonged exposure of the mammary glands to progesterone promotes a more extensive anatomic development of the secretory alveoli. While progesterone stimulates anatomic development, it inhibits the functional development of the secretory epithelium. Progesterone also inhibits the production of intracellular enzymes necessary for normal milk secretion. This inhibitory effect of progesterone is lost just prior to parturition, and this is one factor promoting lactogenesis.
Prolactin, a protein hormone, is unique in that its release from the adenohypophysis is primarily regulated by a humoral inhibitor factor from the hypothalamus. in the absence of this inhibitory factor (which is believed to be the catecholamine dopamine), there is a continuous and relatively high rate of prolactin release. in most domestic animals, blood levels of prolactin gradually increase late in gestation, with an abrupt increase at parturition. During late gestation, prolactin receptors in mammary glands also increase under the influence of rising estrogen. Prolactin promotes both anatomic and functional development of the secretory epithelium of mammary glands to promote milk secretion, so the abrupt rise at parturition is appropriate for lactogenesis.
Under the influence of prolactin, secretory cells lining alveoli produce intracellular enzymes necessary for milk secretion. The functional development of alveolar secretory cells is also enhanced by glucocorticoids, which increase in the blood prior to parturition in most species.The placenta of ruminants produces a protein hormone, placental lactogen, or chorionic somatomammotropin, which is similar in structure and function to prolactin. Placental lactogen production increases in late gestation in ruminants and is believed to be more responsible for mammary gland development in these species than prolactin from the adenohypophysis.
Galactogenesis
The continuation of lactation requires stimuli to promote milk production and removal or inhibition of stimuli that retard milk production. stimulation of the nipples (teats) by either milking or suckling elicits an abrupt increase in blood levels of prolactin. The increased secretion of prolactin is the result of a neural reflex mediated through the hypothalamus that regulates prolactin release from the adenohypophysis. The increase in prolactin is relatively short in duration (minutes to an hour). This periodic and relatively brief surge of prolactin is essential to maintain normal lactation in most species, but prolactin does not appear to be an essential regulator of lactation in cows. supplementation with prolactin does not increase milk secretion in cows, and lactation is maintained in cows in spite of severe reductions in blood levels of prolactin.
Growth hormone appears to be especially important in cows, whose blood levels of growth hormone are directly correlated with the maintenance and level of milk production. Growth hormone supplementation increases milk production in cows by 10-40%. Growth hormone supplementation in cattle is associated with a variety of physiologic changes that promote milk production. These include increased mobilization of body energy stores and alterations in overall protein metabolism in other organs to provide substrates for milk secretion, increased food intake, increased nutrient absorption from the gastrointestinal tract, and improved efficiency of the conversion of nutrients to milk by the mammary gland. it is presumed that many of the effects of growth hormone are mediated via insulin-like growth factors (IGFs), whose production is increased by growth hormone (see Chapter 12). Even though growth hormone appears to be a primary regulator of milk production in cows, milking or suckling does not produce an immediate release of growth hormone in the lactating cow.
Routine milking or suckling to remove milk from the mammary glands is essential for continued milk production. When milking is stopped abruptly (probably the best way to dry up a cow), a number of changes in the udder occur. At the end of 24 hours, the alveoli become distended to a maximum, and the capillaries are full of blood. Between 36 and 48 hours, there is a decrease in the number of patent (open) capillaries, and the alveoli do not respond to intravenous oxytocin. A protein that inhibits milk production, feedback inhibitor of lactation (FIL), is apparently produced in the mammary gland as milk is produced. This protein has a local effect to inhibit milk production. Other components of milk may have similar inhibitory effects, and routine removal of these inhibitory factors promotes the continuation of lactation.
Milk Ejection or Letdown
Milking or nursing alone can empty only the cisterns and largest ducts of the udder. in fact, any negative pressure causes the ducts to collapse and prevents emptying of the alveoli and smaller ducts. Thus, the dam must take an active although unconscious part in milking to force milk from the alveoli into the cisterns. This is accomplished by active contraction of the myoepithelial cells surrounding the alveoli (milk ejection, or milk letdown). These myoepithelial cells contract when stimulated by oxytocin, a hormone released from the neurohypophysis of the pituitary as a result of a neuroendocrine reflex. The afferent side of the reflex consists of sensory nerves from the mammary glands, particularly the nipples or teats. Afferent information reaches the hypothalamus, which regulates the release of oxytocin from the neurohypophysis (see Chapter 12). Suckling the teats by the young is the usual stimulus for the milk ejection reflex, but whether milk is withdrawn from the teat or not, the milk ejection reflex produces a measurable increase in the pressure of milk within the cisterns of the udder.
The milk ejection reflex can be conditioned to stimuli associated with milking routine, such as feeding, barn noises, and the sight of the calf. it can also be inhibited by emotionally disturbing stimuli, such as dogs barking, other loud and unusual noises, excess muscular activity, and pain. stressful stimuli increase the activity of the sympathetic nervous system, which can inhibit the milk ejection reflex. This inhibition occurs both at the level of the hypothalamus via an inhibition of oxytocin release and at the level of the mammary gland, where sympathetic stimulation can reduce blood flow and directly counteract the effect of oxytocin on myoepithelial cells.
oxytocin release typically occurs as a surge within a minute or two after initiation of the reflex by some tactile or environmental stimulus, and the plasma half-life of oxytocin (a small peptide hormone) is but a few minutes. Hence, milking or suckling should begin in close association with stimuli known to stimulate oxytocin release, such as washing the udder and stimulation of the teats. if failure to get an adequate stimulus for milk ejection, possibly because of inadequate preparation before milking, becomes habitual, the lactation period may be shortened by excessive retention of milk in the udder.
Essentially all the milk obtained at any one milking is present in the mammary gland at the beginning of milking or nursing. However, milking does not usually remove all of the milk in the gland. Up to 25% of the milk in a gland usually remains after milking. some of this residual milk can be removed after injections of oxytocin, but the routine use of such injections tends to shorten the lactation period.
Colostrum
Colostrum, the first milk produced upon delivery of the newborn, is important for the survival and vitality of newborn domestic animals. one of the unique differences between colostrum and typical milk is that colostrum contains a high concentration of immunoglobulins produced by the immune system of the dam. These immunoglobulins are concentrated in the milk by selective transport by the epithelial cells lining the alveoli and are needed by the neonate to provide temporary immune protection against infectious agents in the environment.
During the first day or two of life, most domestic neonates can absorb intact immunoglobulins from their gastrointestinal tract into the blood. After this period, immunoglobulins cannot be absorbed intact, and consumed immunoglobulins are digested in a manner similar to other ingested proteins. Closure is the term given to the changes that occur in the gastrointestinal tract after the first or second day of life that prevent the continued absorption of intact immunoglobulins. Colostrum consumption is especially important in domestic farm animals because of limited transfer of immunoglobulins from the dam to the fetus via the placenta. in some mammals, including humans, transfer of immunoglobulins via the placenta occurs to a greater extent, so consumption of colostrum by neonates is less important.
Colostrum is also a source of energy for the newborn, since most are born with limited amounts of body fat and other sources of metabolic energy. The primary sources of energy in colostrum are milk proteins and lipids, because colostrum has a relatively low concentration of lactose. Recall that progesterone inhibits development of enzymes necessary for lactose synthesis until just prior to parturition. The colostrum of most species also has tends to have relatively high concentrations of vitamins A and D and iron, but some species differences in composition do exist.
Cessation of Lactation
Daily milk production reaches a maximal value at some point after lactation begins and then gradually declines over time in most species. The decline in milk production is associated with a gradual decrease in the number of active alveoli and an increase in the relative amount of connective tissue. Mammary gland involution is the term describing the conversion of a milksecreting gland with milk-filled alveoli to one characterized by small, nonsecreting alveoli surrounded by an extensive amount of connective tissue. Extending the period of lactation is economically important for some species (e.g., dairy cows), but not all (e.g., piglets may be weaned prior to peak milk production by sows).