Seminiferous Tubules and Spermatogenesis
Seminiferous Tubules
The epithelium lining the seminiferous tubules contains two cell types, the sustentacular, or Sertoli cells, and the developing spermatozoa and their precursor germ cells (Fig.
25-1).The large sustentacular cells extend from the base of the epithelium to the lumen of the seminiferous tubules. Their shape is highly irregular because they encircle the developing germ cells (Fig. 25-1). Sustentacular cells secrete a fluid that bathes the developing germ cells and assists with the transport of spermatozoa from the tubules into the rete testis after their release from the sustentacular cells. One component of this fluid, androgen-binding protein, transports androgens from their site of synthesis in the testis to the epididmyis, where they are required for maturation of the spermatozoa. Interstitial cells (Leydig cells) in the connective tissue between seminiferous tubules secrete testosterone, the primary androgen originating in the testicles.
The composition of the interstitial fluid between sustentacular cells and within the lumen of the seminiferous tubules differs from typical interstital fluid. The differences are due in part to a selectively permeable barrier (blood— testis barrier) between these fluids and fluid
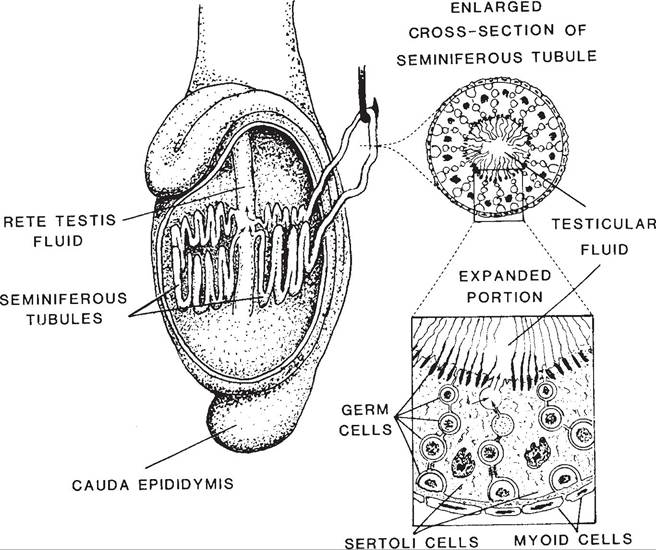
Figure 25-1. Location and microscopic anatomy of seminiferous tubules. Note the irregular border of the sustentacular (Sertoli) cells and their relation to the developing germ cells. (Reprinted with permission of Wiley- Blackwell from Hafez E.S.E. and Hafez B. Reproduction in Farm Animals. 7th ed. Philadelphia: Lippincott Williams & Wilkins, 2000.)
outside the tubules. The cell junctions between adjacent sustentacular cells and between myoid cells that surround the tubules contribute to this barrier (Fig.
25-1). The luminal concentrations of cellular secretions, such as androgenbinding proteins and androgens, are higher than in typical extracellular fluid because this functional barrier sequesters them.Germ Cells and Spermatogenesis
Spermatogenesis is the term for all processes involved in the formation of mature male gametes from the most undifferentiated germ cells. it includes several mitotic cell divisions followed by two meiotic cell divisions, during which the chromosome number is reduced from diploid to haploid. This series of cell divisions is termed Spermatocytogenesis (Fig. 25-2).
Some of the cells resulting from the mitotic cell divisions of the most undifferentiated germ cells remain at the base of the epithelium to maintain the supply of stem cells. others begin the sequence of cell divisions (mitotic followed by meiotic divisions) and developmental changes to become spermatozoa. The mitotic cell divisions double the number of cells at each step, so a single spermatogonium gives rise to many spermatozoa.
Meiosis entails two cell divisions and occurs only during the development of gametes in the testis and ovary. Prior to the first division, the DNA is replicated in a manner similar to that of mitotic cell division (see Chapter 2). This replication results in chromosomes that consist of two identical chromatids. In preparation for the first meiotic division, homologous chromosomes pair up along the middle of the cell. (Homologous chromosomes are the similar
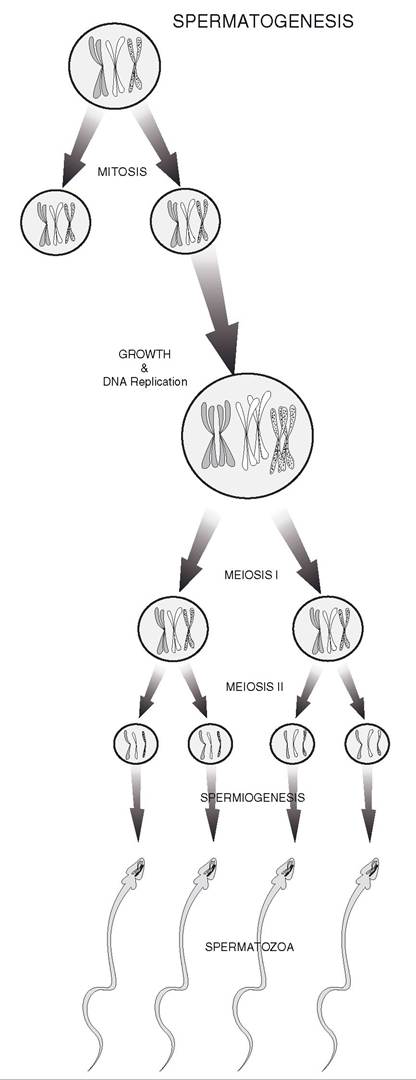
Figure 25-2. Spermatogenesis: cell division and structural changes resulting in the formation of spermatozoa.
chromosomes of a typical pair, each of which was contributed by one of the parent animals.) During the first meiotic division, one chromosome of each homologous pair moves into each daughter cell. Which individual chromosome of the homologous pairs moves to which daughter cell appears to be random.
This mixing among homologous pairs provides for genetic variation among the offspring. After the first division, each daughter cell has a haploid number of chromosomes, but each chromosome consists of two chromatids. During the second meiotic division of the two daughter cells, each of the resulting four cells receives one of the chromatids. The overall result of meiosis is the production of four daughter cells, each of which has a haploid number of chromosomes (Fig. 25-3).in mammals, two of the daughter cells contain the Y chromosome that produce a male offspring (XY) when combined with an X
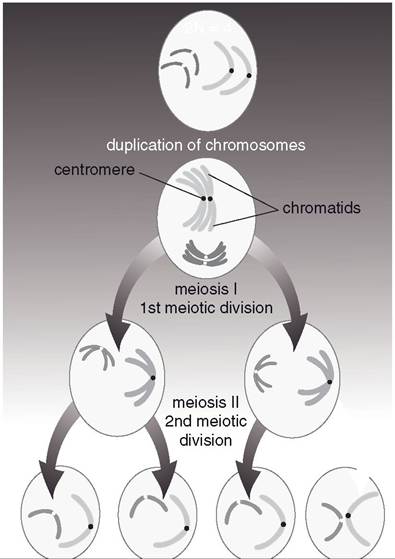
Figure 25-3. Meiosis. Two cell divisions resulting in four cells with haploid number of chromosomes.
chromosome from an ovum. The other two contain the X chromosome to produce female offspring (XX) when united with the X chromosome-containing ovum. However, in avian species, all spermatozoa contain the same sex chromosome, known in these species as the Z chromosome. Female birds carry one Z and a second sex chromosome, the W chromosome. An ovum may contain either a W or Z chromosome, so the ovum is the gamete that determines the sex of the offspring in birds.
When homologous chromosomes are paired in preparation for the first meiotic division, crossing over may occur. During this process, similar regions of chromosomes may be exchanged between homologous chromosomes. such exchanges further increase genetic variability among the offspring, for they may now inherit chromosomes that are different from either parent chromosome.
Spermatid is the term for the cells resulting from the second meiotic division in the seminiferous tubules. spermatids undergo a series of functional and structural changes to become spermatozoa, and this process is termed spermiogenesis (Fig. 25-2). There is no further cellular division after second meiotic division, so there is no further increase in cell number after that division.
Spermatozoa Morphology and Spermatogenesis
When first produced, spermatids are round immobile cells, while fully formed spermatozoa are mobile and consist of a head and tail (Fig. 25-4). The head contains the nucleus, primarily consisting of condensed genetic material (DNA), and an acrosome, a membranous sac that lies immediately under the plasma membrane at the tip of the head and extends down over the nucleus (Fig. 25-5). The tail has a central core of microtubules and filaments that provide motility. The middle piece of the tail contains a dense collection of mitochondria that provide energy for spermatozoan motility.
The acrosome is essentially a membranous sac of hydrolytic enzymes, including acrosin
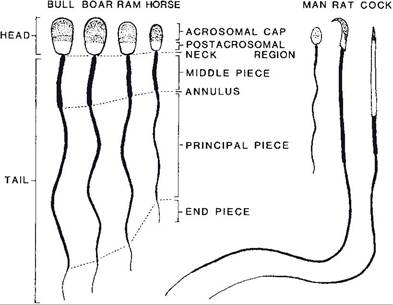
Figure 25-4. Comparison of the spermatozoa of farm animals and other vertebrates. (Reprinted with permission of Wiley-Blackwell from Hafez E.S.E. and Hafez B. Reproduction in Farm Animals. 7th ed. Philadelphia: Lippincott Williams & Wilkins, 2000.)
and hyaluronidase. some of these enzymes are released from the acrosome during fertilization and facilitate the fusion of the male and female gametes. The release and exposure of these enzymes during fertilization is termed the acrosome reaction.
The conversion of the round spermatid to the elongated spermatozoon with its acrosome containing head and tail requires reshaping of the original spermatid, synthesis and packaging of acrosomal enzymes, organization of microtubules, and elimination of excess cytoplasm and cell membrane. The sustentacular cells, in which the developing spermatozoa are embedded, assist with some of these conversions. For example, sustentacular cells phagocytose residual bodies of excess cytoplasm and membrane after the release of fully formed spermatozoa.
Rates and Timing of Spermatogenesis
Daily production of spermatozoa has been estimated as 4.4 ? 109 in the ram and 2 ? 109 in the bull.
Eight ejaculations of the bull within an hour reduced the semen from 4.2 mL at the first collection to 2.9 mL at the eighth collection, and the number of spermatozoa was correspondingly reduced from 1.7 billion to 98 million per milliliter. These data indicate that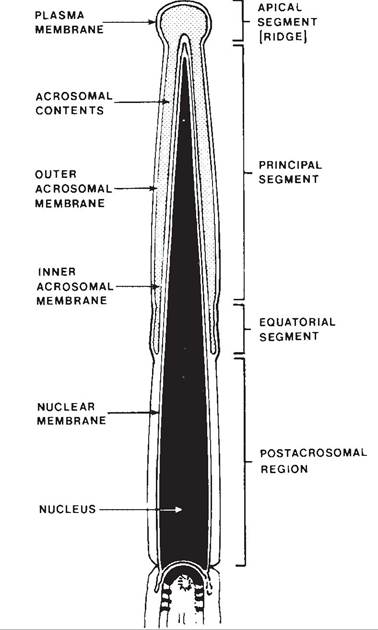
Figure 25-5. A sagittal section of a bovine sperm head showing the various anatomic subdivisions. (Reprinted with permission of Wiley-Blackwell from Hafez E.S.E. and Hafez B. Reproduction in Farm Animals. 7th ed. Philadelphia: Lippincott Williams & Wilkins, 2000.)
normal animals produce adequate spermatozoa even when used often for breeding. However, these data also suggest that several days of sexual rest may increase numbers of spermatozoa in animals whose numbers are abnormally low. A correlation between daily production and testicular size has been found for several species.
The time required for spermatogenesis (from spermatogonium to fully formed and released spermatozoa) varies with species, but it is a matter of weeks to months rather than days. in most farm animals, this period is about 2 months.