SPERMATOGENESIS
1. Define spermatogenesis.
2. Spermatids undergo nuclear.and cytoplasmic changes and develop a tail. What is this maturation phase called?
3. What is spermiation?
4. Where is the fertilizing ability of spermatozoa attained? Where are they stored? What happens to spermatozoa that are not ejaculated?
5.
What function is served by the spermatogenic wave?6. Describe the negative feedback system that relates to the production of testosterone by Leydig cells. Why is luteinizing hormone called interstitial cell stimulating hormone (ICSH)?
7. What is the role of testosterone in spermatogenesis?
8. What are the assumed roles of FSH in the male?
9. Aside from spermatogenesis, what are other functions of testosterone in the male?
0. What embryonic structures stimulated by testosterone become tubular portions of the male reproductive system?
11. What metabolic function is served by testosterone?
2. What are C-16 unsaturated androgens that are secreted by boar testes?
The term spermatogenesis refers to the entire process involved in the transformation of germinal epithelial cells (stem cells) to spermatozoa and can be divided into two phases: spermatocytogenesis and spermiogenesis. Spermatocytogenesis is the proliferative phase whereby spermatogonial cells multiply by a series of mitotic divisions followed by the meiotic divisions that produce the haploid (n) number of chromosomes (Figure 14-15).
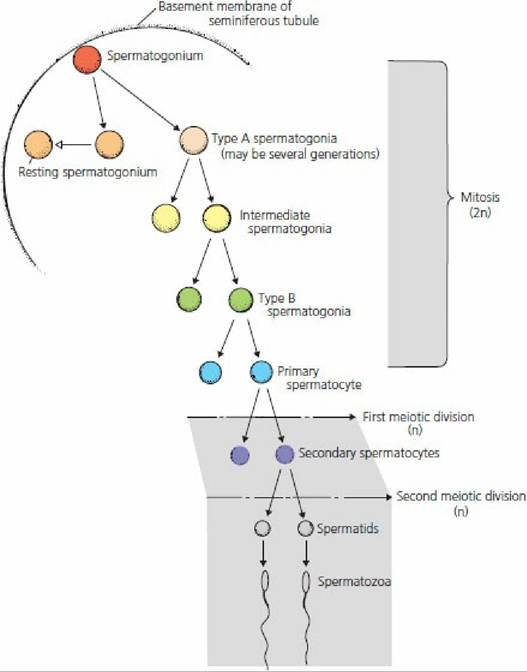
■ FIGURE 14-15 Diagrammatic representation of the stages of spermatogenesis in mammals. The chromosome number (2n, diploid; n, haploid) is also shown for each stage. (From Pineda MH. The biology of sex. In: Pineda MH, Dooley MP, eds. Veterinary Endocrinology and Reproduction. 5th edn. Ames, IA: Iowa State Press, 2003.)
The stem cells (spermatogonia) are located in the basal compartment of the seminiferous tubules (see Figure 14-4).
The mitotic division of a spermatogonium results in one cell being a replacement for the cell that has just divided (it stays in the basal compartment). The other cell becomes a type A spermatogonium, which migrates through the Sertoli cell barrier to the adluminal compartment. Type A spermatogonia undergo mitotic division (sometimes involving several generations) until large numbers (variable among species) of type B spermatogonia have been produced. Type B spermatogonia undergo the last of the mitotic divisions, which results in the formation of primary spermatocytes with 2n chromosome numbers. Primary spermatocytes undergo meiotic division (described previously) to form secondary spermatocytes, which in turn undergo meiotic division to form spermatids (n chromosome numbers). In the bull, 64 spermatids are formed from one type A spermatogonium.The second phase of spermatogenesis, spermiogenesis, involves maturation of the spermatids while they are still in the adluminal compartment. Spermiogenesis comprises a series of nuclear and cytoplasmic changes and transformation from a nonmotile cell (not able to move) to a potentially motile cell in which a flagellum (tail) has formed. The mature spermatids produced during the final phase of spermiogenesis are released into the lumen of the seminiferous tubules as spermatozoa.
The release of matured spermatids into the lumen of the seminiferous tubules is known as spermiation. Spermatozoa from several animal species are compared in Figure 14-16.
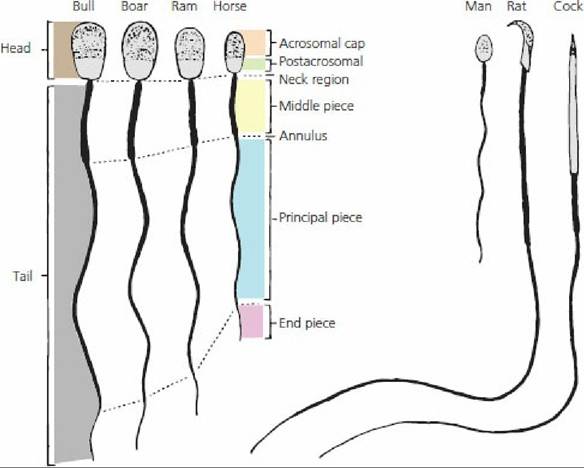
■ FIGURE 14-16 Comparison of the spermatozoa of farm animals and other vertebrates. The major structural features are given. Note the differences in the relative size and shape. (From Hafez ESE, Hafez B. Reproduction in Farm Animals. 7th edn. Baltimore, MD: Lippincott Williams & Wilkins, 2000.)
Epididymal Transport
The newly formed spermatozoa are essentially immotile.
They are transported to the epididymis by fluid secretions into the seminiferous tubules and rete testis and by activity of contractile elements in the testis that direct fluid flow to the head of the epididymis.The fertilizing ability of an animal is attained progressively during the transit of spermatozoa through the epididymis. Changes include development of unidirectional (as opposed to circular) motility, changes in nuclear chromatin (DNA-protein complex), and changes in the nature of the surface of the plasma membrane.
The major site of sperm storage within the male reproductive tract is the tail (last portion) of the epididymis. About 70% of the total number of spermatozoa in the ducts outside the rete testis (excurrent duct system) are found in the tail of the epididymis.
Many of the spermatozoa formed in the testes are either phagocytized in the excurrent duct system or lost into the urine. About 85% of the daily sperm production in sexually inactive rams are voided in.the urine.
Spermatogenic Wave
If all segments of the seminiferous tubules were involved in the same activity at the same time, a continuous supply of spermatozoa would not be produced because about 64 days (in the bull) are required in the adluminal compartment. While this development is proceeding, a new type A spermatogonium migrates through the Sertoli cell barrier into the adluminal compartment to begin its development behind the developing type A spermatogonium that preceded it. In the bull, this occurs every 14 days. Since 64 days are required for development to spermatozoa, there will be 4.6 cycles (64/14) of beginning development before the first cycle from a given area of seminiferous epithelium begins to arrive at the rete testis. A cycle is defined as a series of changes in a given area of seminiferous epithelium between two appearances of developmental stages. A portion of tubule at one stage is usually adjacent to portions of tubule in stages just preceding it or following it in time.
This sequential change in the stage of cycle along the length of the tubule is known as the spermatogenic wave. A spermatogenic wave that involves a 12-day cycle is illustrated in Figure 1417. The wave involves a sequence of stages beginning with the less advanced stages in the middle of the loop to progressively more advanced stages nearer the rete testis. The stages proceed in opposite directions from the site of reversal at the middle of the loop towards the rete testis.
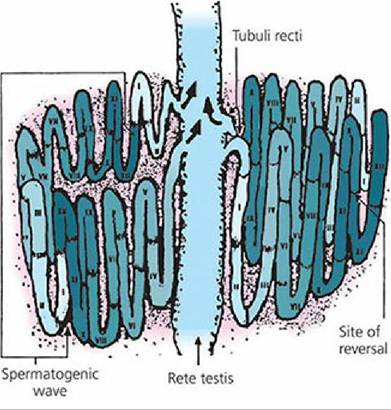
■ FIGURE 14-17 A seminiferous tubule in which the wave of the seminiferous epithelium is schematically represented along the length of the tubule. The succession of stages I to XII (a 12-day cycle), the site of reversal in the middle of the tubule, and the relationship of the wave to the rete testis are shown. The more advanced stages of each wave are located nearer the rete testis. Any one tubule may have as many as 15 spermatogenic waves. (From Hafez ESE, Hafez B. Reproduction in Farm Animals. 7th edn. Baltimore, MD: Lippincott Williams & Wilkins, 2000.)
A large number of spermatozoa are produced daily in the normal male animal, about 6.0 ? 109 spermatozoa in the bull and about 16.5 ? 109 in the boar. In the bull, daily sperm production increases with age, reaching a maximum at about 7 years.
Hormonal Control
Leydig and Sertoli cells are responsible for hormone production within the testes. The production of testosterone by Leydig cells is controlled by the gonadotropin known as luteinizing hormone (LH) (sometimes called interstitial cell stimulating hormone, or ICSH). Low levels of testosterone cause an increase in LH secretion by the anterior pituitary. The increase in LH secretion causes the Leydig cells in the testes to secrete testosterone; when increased, testosterone inhibits the further secretion of LH and testosterone levels are thus stabilized.
A subsequent decline in testosterone again stimulates LH secretion and the cycle is repeated; this is known as a negative feedback system.The influence of testosterone on spermatogenesis requires it to diffuse from the interstitial tissues into the seminiferous tubules. Within the seminiferous tubules, it seems that testosterone maintains spermatogenesis by supporting the meiotic process.
Another gonadotropic hormone, follicle stimulating hormone (FSH) from the anterior pituitary, stimulates production of an androgen-binding protein by the Sertoli cells. Androgen-binding protein is secreted into the lumen of the seminiferous tubules and binds with testosterone and other androgens to stabilize their concentrations and ensure appropriate amounts for spermatogenesis. It is also believed that FSH stimulates the secretion of estrogens by the Sertoli cells. The actual secretion of estrogen might arise from the intracellular conversion of testosterone (originating from Leydig cells) by the Sertoli cells. The Sertoli cells are also the source of a hormone known as inhibin, which inhibits secretion of FSH by the anterior pituitary.
Whereas LH is required continuously.for spermatogenesis (testosterone-supported meiosis), FSH is not essential for the maintenance of spermatogenesis once it has been initiated. Initiation of spermatogenesis at puberty and after physiologic or pathologic interruptions requires FSH.
Other Functions of Testosterone
In addition to its spermatogenic activity, testosterone fulfills other functions in the peripheral circulation. After secretion of testosterone by Leydig cells into the interstitial space of the testes, a greater amount diffuses into the blood and lymphatic capillaries than that which diffuses into the seminiferous tubules. After entrance into the blood, testosterone is bound loosely with a plasma protein for its transport. Within 15 to 30 minutes, the testosterone is released from the protein to be fixed to target tissues or to be degraded, mainly by the liver, into inactive products that are subsequently excreted.
Other functions of testosterone include the development and maintenance of libido, secretory activity of the accessory sex glands, and general body features associated with the male.
Libido refers to sexual drive. It can be effectively eliminated by castration (removal of the testes). Castrated animals usually, but not invariably, lack libido. Small amounts of testosterone from other sources such as the adrenal gland (interconversion potential) might be sufficient to provide libido in some animals.
The structural development and physiologic functioning (production of secretions) of the accessory sex glands are influenced by testosterone. In this regard, hyperactive prostate glands (enlargement) can be treated effectively by estrogen administration. The estrogen inhibits the secretion of LH, and testosterone production by the Leydig cells is suppressed. A reduced concentration of testosterone causes the hyperactive prostate gland to reduce its activity and its size decreases.
General body features associated with the male (secondary sexual characteristics) are influenced by testosterone. These features include increased bone growth (heavier bones), greater muscling, thicker skin, and deeper voice (in the bull). During fetal growth, testosterone directs the descent of the testes. The presence or absence of testosterone determines the respective development of a penis and scrotum or a clitoris and vagina. Before sexual differentiation in the embryo, the structures needed for the development of either sex are present. With normal male hormonal stimulation, the Wolffian (mesonephric) ducts become tubular portions of the male reproductive system and the Mullerian (paramesonephric) ducts regress. In the female the Mullerian ducts become tubular portions of the reproductive system and the Wolffian ducts regress.
Metabolically, testosterone has protein anabolic functions that affect the greater muscling potential of males. It is probable that the thicker skin and laryngeal changes of the male are also related to this function of testosterone. Because of the desirability for more muscle and less fat in meat-producing animals, there has been a tendency to use noncastrated males for meat production. The protein anabolic effect obtained from testicular testosterone would thereby be retained.
Other Androgens
Testosterone is one of several steroid hormones classified as androgens. In addition to testosterone, the boar testes secrete large amounts of compounds known as C-16 unsaturated androgens. These androgens act as pheromones when they are excreted in boar saliva and they cause the sow in heat to adopt the mating posture. When the C-16 unsaturated androgens are excreted in urine, they contribute to the characteristic odor of boar urine. These compounds are also responsible for the undesirable flavor of boar meat, which is known as boar taint.
■