Biology and Epidemiology of Epizootic Emmonsia, Blastomyces and Emergomyces Species
In their mycelial form, many members of Ajellomycetaceae resemble each other in the production of spherical or disc-shaped conidia borne on narrow, cylindrical or swollen stalks (Dukik et al.
2017; Fig. 7.2). However, in mammalian hosts, the genera Emmonsia, Blastomyces and Emergomyces can be differentiated by the appearance of their thermodependent growth phase. Blastomyces parvus produces uninucleate, typically 10-40 μm wide, thin-walled cells (Sigler 2005), whereas cells of A. crescens (adiaspores) are multinucleate and up to 400 μm in size (Dvorak et al. 1973; Boisseau-Lebreuil 1975; Sigler 2005). Moreover, there are differences between these species in the temperatures at which thermal dimorphism occurs, with A. crescens and B. parvus switching at temperatures approaching 37 and 40 °C,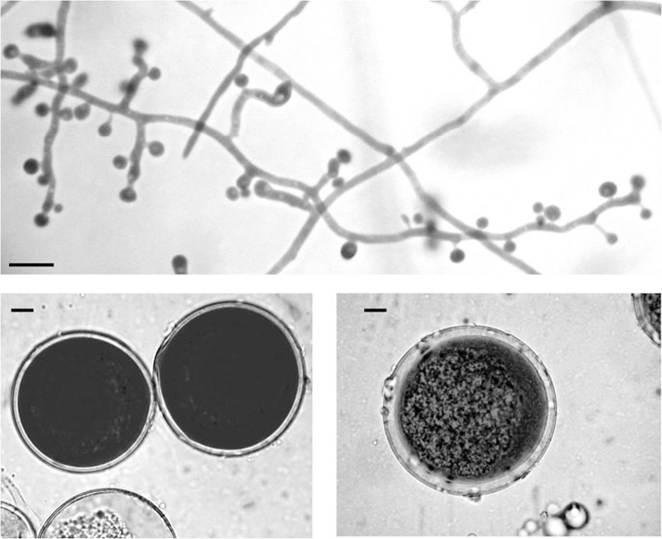
Fig. 7.2 Microscopic appearance of Emmonsia crescens, NCPF 4268 on Sabouraud's agar after 3 weeks at 30 °C (Top), 3 weeks at 37 °C (bottom left) or blood agar after 3 weeks at 37 °C (bottom right). Scale bar = 10 μm
respectively (Borman et al. 2009; Jiang et al. 2018). Similar adiaspores can easily be produced from mycelial cultures of A. crescens in vitro if cultures are moved to and maintained at 37 °C, with typical thick-walled multinucleate cells developing after several weeks (Fig. 7.2).
Judging from historical reports in which adiaspore size was accurately recorded, A. crescens was encountered worldwide, and apparently has a broad host range, although it has a focus with small terrestrial animals. Infections have been reported in foxes, Eurasian otters, stoats, weasels, and various species of mice, voles and shrews in Central and Eastern Europe (Sharapov 1969; Krivanec 1977; Krivanec and Otcenasek 1977; Hubalek 1999; Borman et al.
2009), ferrets in New Zealand (Lugton et al. 1997), hairy nosed wombats in Australia (Mason and Gauhwin 1982), striped skunks and ground squirrels in Canada (Leighton and Wobeser 1978; Albassam et al. 1986), hedgehogs, moles, voles and shrews in France (Doby et al. 1971), otters, voles, muskrats and lemmings in Finland (Jellison et al. 1960), voles, shrews and otters in Sweden (Jellison 1969), mole-rats in Zambia and Israel (Hubalek et al. 2005) and various free-living mammals in the UK (McDiarmid and Austwick 1954; Tevis 1956; Austwick 1968, Chantrey et al. 2006; Borman et al.2009). Indeed, previous reviews have enumerated in excess of 100 host animal species for A. crescens (Sigler 2005). Conversely, B. parvus has a lower prevalence, with reports from the USA, Kenya, Zambia, Israel, Australia and Eastern Europe, and with a narrower host range (Jellison 1969; Krivanec et al. 1976; Krivanec 1977; Mason and Gauhwin 1982; Hubalek et al. 2005; Sigler 2005).
Although the natural habitat of Emmonsia-like species has not been precisely defined, there is strong evidence that infection is acquired by inhalation of conidia from soil and/or nesting materials of burrowing mammals. Emergomyces africanus has been detected in soil and air samples from South Africa (Schwartz et al. 2018a, b), but natural infections of animals have not been proven (Cronje et al. 2017). Experimental infections have successfully been induced in a variety of mammalian species after intranasal inoculation of conidial suspensions prepared from the mycelial phase of A. crescens (Jellison 1969), with mature adiaspores measuring up to 100 μm developing within 15 days of inoculation. Moreover, several reports have detailed the direct isolation of Emmonsia-like fungi from soil and nest materials with or without passage through rodents (Jellison 1969; Hubalek et al. 1995; Peterson and Sigler 1998; Sigler and Flis 1998 and later editions), and viable adiaspores have been recovered from the digestive tracts of rodents and also from the dung of larger carnivores that prey on infected rodents (Krivanec et al.
1975; reviewed in Sigler 2005). Finally, studies from Czechia reported high prevalence in nesting rodents in wooded areas (Hubalek et al. 1995), but failed to find evidence of adiaspiromycosis in rodents that do not nest in soil (Hubalek 1999). However, although the above data are good evidence that A. crescens may be vectored by animals (via predation/ excretion/decomposition), there exists no evidence that the pathogen can be transmitted directly from animals to man.Several studies have evaluated prevalence of infection. Studies from Czechia, France and UK estimated prevalence rates between 30 and 70% (e.g. Dvorak et al. 1973; Hubalek 1999; Borman et al. 2009), with higher rates in rodents than in carnivores and a predilection for mustelid carnivores (Doby et al. 1971; Krivanec et al. 1975; Borman et al. 2009). Several reports also suggested seasonal variation in infection rates (Dvorak et al. 1969; Hubalek et al. 1993), and presented evidence for wide heterogeneity of adiaspore sizes within the lungs of single affected mammals, suggestive of multiple exposures, possibly by several species. Similarly, the analysis of infectious burdens in several studies revealed large variations in mean adiaspore numbers amongst infected animals, with heavy infections associated with poor general condition, emaciation and even death (Jellison 1969; Hubalek et al. 1995; Simpson and Gavier-Wilden 2000; Borman et al. 2009). Finally, isolated cases of adiaspiromycosis have been sporadically reported in domestic mammals, including farm animals, dogs, goats and horses (reviewed in Sigler 2005).
7.3