Brief History of an Emerging Disease
When “white-nose syndrome” (WNS) emerged, nobody anticipated that it would be the start of one of the most devastating mass mortalities in mammals ever recorded. In 2006 a group of hibernating little brown bats (Myotis Iucifugus) was photographed in Howes Cave, New York State, USA, depicting the animals with distinct white powdery patches around their muzzles and on their wing membranes, name-giving for WNS.
During the following winters of 2006-2008, it was noticed that bats displayed aberrant hibernation behaviour in the vicinity of several cavernous hibernacula like day-flight or aggregation of bats close to the cavern entries. In mid-March 2007, a tenfold increase of midday flight compared to the previous 25-year record high was reported (Blehert et al. 2009). But the most dramatic observation was a decline of up to 75% of hibernating bats in this region at that time. A fevered search for the causative agent followed, and several laboratories tried to identify a possible microbial agent or toxic component. Several bat carcasses underwent pathological investigations including necropsy and histopathology. One of the obstacles during the search was that during shipment from the field to the investigation centres, the white powdery substance seen on the affected bats rendered invisible (Meteyer et al. 2009), and it took many attempts until finally a fungal organism was consistently cultured and identified. In late 2008, Blehert et al. described a novel fungus closely related to the psychrophilic and keratinophilic species Geomyces pannorum. But because of distinct, characteristically formed conidia as well as differences in the ITS genes, the isolated fungus was identified as a new species: Geomyces destructans (Blehert et al. 2009; Gargas et al. 2009). Meanwhile no other microbial agent or toxic component was consistently found in all investigated bats, making G. destructans the likely cause of disease. A few years later, Koch's postulates were fulfilled in experimental infection trials proving the fungus as the causative agent (Lorch et al. 2011). Soon it became evident that the fatalities associated with the fungus were rapidly spreading in a radial fashion away from the index cave to further wintering sites. Such pattern is well known as a characteristic feature of pathogens introduced into a naive population. Today, 10 years after the first detection, more than six million hibernating bats succumbed to the infection (US Fish and Wildlife Service March 2016a) grimly making earlier modelling studies, that predicted regional extinction of the once abundant little brown bat within 16 years, become true (Frick et al. 2010). Seven North American bat species are affected and hibernacula positive for G. destructans are currently found in 26 US states and 5 Canadian provinces (Hayman et al. 2016), and it is likely to proceed further (Fig. 13.1). In 2013, improved phylogenetic techniques indicated that the former classification of the fungus needed to be reclassified as Pseudogymnoascus destructans (Minnis and Lindner 2013).While these dramatic events enfolded in North America, Europe feared similar consequences if the fungus should enter European hibernacula. Hence, it was rather surprising when it became evident that P. destructans is enzootic in Europe and hibernating bats in various European countries carry the fungus (Fig. 13.2) (Puechmaille et al. 2011; Zukal et al. 2016). However, in Europe, P. destructans
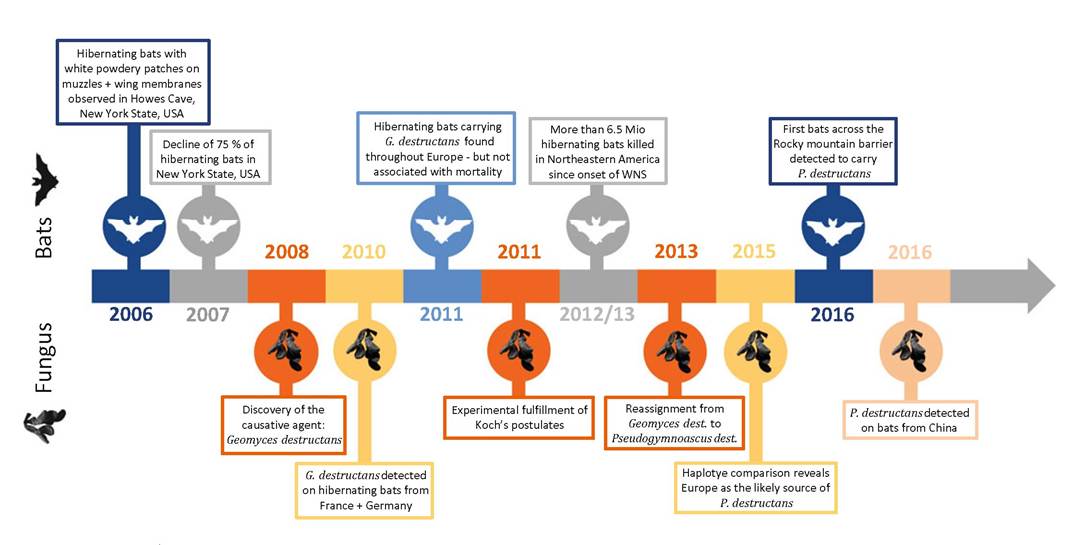
Fig. 13.1 Timeline of Pseiidogynmoasciis destructans detection and subsequent developments
13 White-Nose Syndrome in Hibernating Bats
Fig. 13.2 An European greater mouse-eared bat (Myotis myotis) carrying white fungal patches of Pseudogymnoascus destructans around its nose, the ears and forearms (Courtesy of C.
Jungmann)
infection is not associated with mass mortalities (Wibbelt 2015). Recently P. destructans was also discovered on four different hibernating bat species in Northeastern China, similar to the situation in Europe without causing fatalities, but rather indicating host resistance to the pathogen (Hoyt et al. 2016). Meanwhile, also in North America, little brown bats were found which recovered from the disease, and during summer bats do clear the infection entirely from their body (Meteyer et al. 2011).
So far hibernating bats are the only host species for which pathogenicity of P. destructans was shown. The slow growth and psychrophilic nature of the fungus make these animals an ideal substrate as the bats' body temperature decreases to almost ambient temperature of the cavernous wintering sites and during their torpor phases of 12-14 days each, the animals remain motionless and leave the fungus undisturbed. During their active life, their body temperature rises like in all warmblooded animals creating an inhospitable environment for the fungus. With the differences between North America and Europe/Asia, the use of the term WNS remains slightly confusing. At the time when the name developed, it referred to a true syndrome: distinct clinical signs leading to (mass) mortalities with unknown aetiology. Although today the causative agent and infection patterns are identified, we are still faced with two different scenarios: hibernating bats infected with P. destructans and ongoing mass mortalities in North America and hibernating bats infected with P. destructans not associated with fatalities in Europe and Palearctic Asia. The indiscriminate use of the term WNS for either situation as well as for bats in North American recovering from WNS might blur future epidemiologic approaches, where a distinction between the two groups infected and dying versus infected and surviving might be difficult to reconstruct. A sharp distinction between histopathologically proven P. destructans infection associated with various clinical signs, including mortality equalling WNS (in North America), and confirmed P. destructans infection of otherwise unremarkable (European) bats equalling P. destructans disease would help to keep a clear line between these important epidemiologic features.
13.2