The Causative Agent
13.2.1 Classification
To date the WNS fungus is classified as Pseudogymnoascus destructans. The genus Pseudogymnoascus is a member of the family Pseudeurotiaceae within the class Leotiomycetes of the phylum Ascomycota.
First classifications after the fungus' detection based on rRNA investigations compared the internal transcribed spacer (ITS) and small subunit (SSU) gene sequences and placed the new fungus in the genus Geomyces (Blehert et al. 2009; Gargas et al. 2009; Chaturvedi et al. 2010), but parsimonious trees for ITS and SSU already indicated a close relationship with Pseudogymnoascus. Lorch et al. (2010) designed primers which selectively amplified DNA targeting a conserved 1506 bp intron and ITS sequences. Soon investigations on soil samples from North American bat hibernacula not only revealed a huge diversity of so far unknown Geomyces species but also showed cross-reactivity with primers thought to be selective for G. destructans (Lindner et al. 2011). Faced by the large number of different Geomyces species and allies in cave soil and a missing taxonomic lead, Minnis and Lindner (2013) employed not only ITS but also nuclear large subunit (LSU) rDNA, DNA replication licensing factor MCM7, RNA polymerase II second largest subunit (RPB2) and translation elongation factor EF-1α (TEF1) to investigate the taxonomic relationship of the various fungal species detected in American hibernacula. The result of this work was a shift of the genera Geomyces, Gymnostellatospora and Pseudogymnoascus from the historically assigned family Myxotrichaceae to the family Pseudeurotiaceae (Minnis and Lindner 2013). At the same time, it was shown that G. destructans was indeed member of the genus Pseudogymnoascus and not Geomyces and was subsequently renamed in Pseudogymnoascus destructans (Minnis and Lindner 2013).
13.2.2 CultureConditions
Pseudogymnoascus destructans is a true psychrophile, slowly growing between 0° and 24 °C.
The temperature optimum lies between 12° and 15 °C with some minor strain-dependent differences, and no growth occurs above 24 °C (Gargas et al. 2009; Verant et al. 2012). There are no specific demands for culture media, but Sabouraud glucose agar containing chloramphenicol and gentamicin seems to be preferred by most studies. However P. destructans grows equally well on corn meal agar, malt extract agar, creatine sucrose agar, yeast and malt extract agar, potato dextrose agar and minimal media agar (Gargas et al. 2009; Chaturvedi et al. 2010; Martfnkova et al. 2010; Puechmaille et al. 2010; Wibbelt et al. 2010; Verant et al. 2012; Lorch et al. 2013a; Khankhet et al. 2014).The type description by Gargas et al. (2009) notes colony diameters after 16 days on corn meal agar with 1 mm at 3.5 °C, 5 mm at 7 °C, 8 mm at 14 °C and no growth at 24 °C. Fungal growth starts with white, smooth-domed colonies, which will
Fig. 13.3 Cultureof
Pseudogymnoascus destructans on Sabouraud glucose agar with excretion droplets

eventually turn into greyish-olive radially from the centre to the outer rim (Gargas et al. 2009; Puechmaille et al. 2010; Verant et al. 2012). However, depending on the isolate, culture medium and temperature, phenotypic differences may occur in growth velocity, exudate production, secretion (Fig. 13.3) and diffusion of soluble pigments and colony reverse colour, the latter ranging from white, grey, grey-green, brown to black (Gargas et al. 2009; Chaturvedi et al. 2010; Verant et al. 2012; Khankhet et al. 2014). Pseudogymnoascus destructans is considered a biosafety level 2 pathogen, and culture and isolation should be performed under the relevant requirements.
13.2.3 MorphologyandReproduction
Gargas et al. (2009) described the fungal morphology for the P. destructans type strain: asymmetrically curved conidia are borne singly at the tips, on the sides or in short chains on verticillately branched conidiophores (Fig.
13.4). Within chains of conidia that undergo rhexolytic dehiscence sometimes, intercalary conidia (arthroconidia) occur with conspicuous separating cells. Conidiophores are erect, hyaline, smooth and thin-walled and narrow (1.5-2 μm wide by 35-90 μm or more in length) and commonly bear verticils of 2-4 branches borne at an acute angle to the stipe. The size of the conidia is 5-12 x 2.0-3.5 μm, tapering basally to 1.5-2.0 μm and apically to 0.5-1.5 μm. They truncate with prominent scars at one or both ends and are smooth and lightly pigmented. Their shape is predominantly curved, sometimes oval, obovoid or cymbiform; at maturity they are moderately thick-walled and readily seceding.However, when P. destructans is grown at temperatures above 12 °C, stress- related changes appear, e.g. hyphae become notably thickened and diffusely septate with differential sequestration of cellular material among segments. Also the conidia change their shape to mostly pyriform or globoid. Increasing temperature seems to result in pronounced deformity of the fungal structures (Verant et al. 2012).
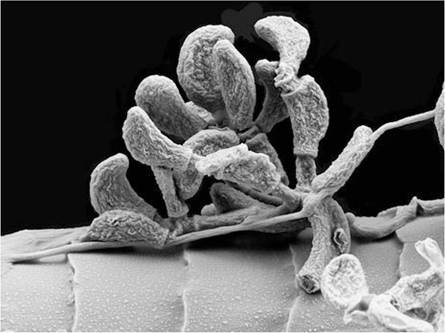
Fig. 13.4 Scanning electron microscopy image of Pseudogymnoascus destructans colonising the hair of a bat
The mating system of P. destructans still remains cryptic, but first molecular investigations of the P. destructans reference strain's (20631-21) genome are suggestive of a heterothallic mating system, because this strain misses the mating type locus MAT1-2-1 high mobility group (HMG) box protein, which in homothallic fungi would be contained in the genome of each isolate. Since P. destructans populations in North America are of single clonal origin (Rajkumar et al. 2011) comprising only a single mating type, isolates of both mating types MAT1-1 and MAT1-2 are coexisting in European hibernacula, indicating that there is the potential for mating populations (Palmer et al.
2014). Examination of mRNA from isolates of these two mating types grown in mixed culture revealed that cocultivation of MAT1- 1 and MAT1-2 strains results in a weak induction of the MAT1-1-3 HMG domaincontaining gene. All these data indicate a heterothallic sexual reproduction pathway (Palmer et al. 2014). Currently in North America, P. destructans spreads by asexual reproduction, as a true mating partner is missing. The introduction of additional isolates could result in sexual reproduction, and possible resulting recombination would allow further adaptation to environment and host species (Palmer et al. 2014).13.2.4 Biochemical Characteristics
Several investigations on the biochemical properties of P. destructans were performed to elucidate potential pathogenicity factors of the fungus. Assays with substrates containing urea agar revealed the presence of urease, while albumin, casein and gelatin showed that the fungus secretes proteinases (Chaturvedi et al. 2010).
Currently the most important enzyme identified in P. destructans is a serine protease called “P. destructans serine protease 1” (PdSP1) (Pannkuk et al. 2015) or “Destructin-1” (O'Donoghue et al. 2015). This subtilisin-like peptidase and its two isoforms PdSP2 and PdSP3 have major proteolytic activities produced by P. destructans, at least under culture conditions. The ability of the fungus to produce proteolytic and hydrolysing enzymes was further tested by API-ZYM tests, and it was found that P. destructans isolates also produce acid phosphatase, alkaline phosphatase, N-acetyl-β-glucosaminidase, β-glucosidase, esterase, esterase lipase, lipase, leucine arylamidase, naphthol-AS-B1-phophohydrolase and valine arylamidase. Enzyme assays on 6% gelatin implicated that lipase and esterase activity are more rapid than proteinase activity, while gelatin degradation was accompanied by alkalinisation (Raudabaugh and Miller 2013).
Substrate suitability assays demonstrated that P. destructans is capable of growth and sporulation on dead fish, insect and mushroom tissues, as well as on media ranging from pH 5 to 11.
It also tolerates media supplemented with 2000 mg/L of calcium and 700 mg/L of sulphur compounds like thiosulphate, L-cysteine or sulphite; thiosulphate is reduced via the generation of hydrogen sulphide gas. The fungus exhibits Class 2 nitrogen utilisation with growth-dependent interactions among different pH and nitrogen sources (Raudabaugh and Miller 2013). Investigations on the influence of the matric potential of substrates used polyethylene glycol for adjustment. It was shown by delayed germination and growth that P. destructans was intolerant to polyethylene glycol-induced matric potential at22.5 MPa and at 25 MPa no germination was at all visible. However, when surface tension of 25 MPa polyethylene glycol medium was decreased, germination and growth of P. destructans were permitted, suggesting a link between substrate suitability and aqueous surface tension altering substances (Raudabaugh and Miller 2013). Pseudogymnoascus destructans is not capable of producing any extracellular biosurfactants or surface tension-reducing compounds, but assays suggest that substrates containing such agents would be beneficial for the growth of the WNS fungus (Raudabaugh and Miller 2013). In a cave environment, surface tensionreducing substrates would contain free fatty acids or lipids that could be hydrolysed to fatty acids by the fungus. Lipids of bat skin and glandular secretions contain free fatty acids (Pannkuk et al. 2013, 2014) making the surface of bat skin an excellent habitat for P. destructans (Raudabaugh and Miller 2013). Members of Pseudoeurotiaceae grow saprophytically on woody tissue and rotting vegetation, and β-glucosidase produced by Pd makes it highly likely that it can degrade cellulosic substrates as long as sufficient moisture will reduce the water potential of debris for P. destructans requirements (Raudabaugh and Miller 2013).
Investigations on the triacylglyceride composition and fatty acyl saturation profiles of P. destructans gave insight into its metabolic capabilities. The fungus produces higher proportions of unsaturated 18 °C fatty acids and triacylglycerides than G. pannorum. At 5 °C it also produces up to a twofold increase in 18:3 fatty acids in comparison to higher culture temperatures, while the triacylglyceride proportion at upper as well as lower temperature growth limits was greater than 50% of total dried mycelia mass (Pannkuk et al. 2014). It is thought that the fungus uses the alteration of acyl lipid unsaturation as a strategy to adapt to cold temperatures as particularly triacylglycerides serve as the main energy reserve in fungi. Moreover, they prevent free fatty acids to reach toxic levels and stimulate reproduction (Brennan and Losel 1978).
13.3