Chikungunya: overview
1.2.1 History and phylogenetics of CHIKV evolution
CHIKV was first isolated and characterized in 1953 during an epidemic of febrile polyarthritis in Tanzania (formerly Tanganyika) [21].
The word “chikungunya” comes from ChiMakonde, the language spoken by the Makonde people, an ethnic group in southeast and northern Mozambique meaning “that which contorts or bends up” and refers to the stooping posture of infected patients due to severe joint pain.Since the 1953 Tanzania outbreak, CHIKV has caused outbreaks in various parts of Africa. The re-emergence of CHIKV epidemic in Africa was documented in 19992000 in Kinshasa where an estimated 50,000 persons were infected. The first documented Asian outbreak took place in 1958 in Bangkok, Thailand. Since then, many outbreaks have been recorded from Cambodia, Vietnam, Laos, Myanmar, Malaysia and Indonesia. There is historical evidence that chikungunya virus originated in Africa and subsequently spread to Asia [22]. Phylogenetic analysis of CHIKV virus sequences originally identified three distinct clades separated primarily by geography designated the West African, CentralZEast African (ECSA) and Asian genotypes. The Asian genotypes have a high degree of sequence identity among themselves whereas the African strains exhibit wider sequence diversity and have been shown to undergo genetic microevolutions even during the course of an epidemic [23]. Recent phylogenetic studies showed that the Indian Ocean and Indian subcontinent outbreaks were caused by virus strains of the Indian Ocean lineage (IOL), which evolved from the ECSA genotype [19, 24]. This lineage first emerged in Kenya in 2004, and subsequently spread to several Indian Ocean islands, India and Southeast Asia. The IOL strains involved in the Indian Ocean and Indian outbreaks possessed the initial adaptive mutation, E1-A226V [25], which is a major genetic determinant of adaptation of CHIKV to Ae.
albopictus vector species and provides a plausible explanation for how this mutant CHIKV caused epidemics in regions lacking the more typical urban vector, Ae. aegypti [26]. Introduction of new viral strains, viz. IOL strains inevitably leads to the question whether particular genotypes of CHIKV are associated with higher virulence or severe disease. In addition, the lack of a suitable animal model for CHIKV makes it difficult to verify such hypotheses [27]. On the other hand, association of the re-emergence of endemic strains with the outbreaks leads to a different question that can be clarified only by a combination of classic epidemiology and comparative genomics: whether the viruses re-emerged due to environmental, population immunity and/or vectorial factors, or whether outbreaks were triggered by adaptive evolution of the virus that endowed it with an increase in fitness and virulence? Therefore, knowledge of the complete genetic blueprint of CHIKV is essential for clarifying these crucial questions.1.2.2 Chikungunya virus (CHIKV):genome structure and organization
Chikungunya is an acute debilitating arthritogenic disease, caused by chikungunya virus (CHIKV) belonging to Alphavirus genus; family Togaviridae, which consists of Alphavirus and Rubivirus genera. Approximately, 40 alphaviruses can infect vertebrates such as humans, rodents, birds, and horses, along with invertebrates. Mosquito vectors are responsible for the transmission between species, thereby rendering alphaviruses to be considered as arboviruses. CHIKV is an enveloped particle and has a single-stranded RNA genome of positive polarity. The genome is approximately 11.8 kb in length [28]. Under electron microscopy in green monkey kidney (Vero) cells CHIKV particles reveal a characteristic Alphavirus morphology.
CHIKV genome consists of polyadenylated RNA and is capped, which encodes two open reading frames (ORFs). Two-third of the genome includes the 5' ORF that encodes four nonstructural proteins, which are involved in genome replication, capping of RNA, polyprotein cleavage, etc., essential for viral replication.
This region is expressed as an nsP1-3 or nsP1-4 polyprotein via cap-dependent translation, which is further cleaved by an nsP2-encoded protease. In context to the structural protein ORF, it is embedded in a subgenomic mRNA, and is translated into proteins via a cap-dependent mechanism. This structural ORF polyprotein is finally cleaved into capsid, envelope glycoproteins E1, and glycoprotein E2. The mature virion comprises 240 heterodimers of E2/E1, which are arranged as trimeric spikes on its surface, and has a diameter of 70 nm (Figure 3). After being translocated by the secretory pathway, these heterodimer spikes penetrate into the plasma membrane of infected cells, and cytoplasmic nucleocapsids containing the genomic RNA and 240 copies of the capsid protein bud from the cell surface for assembly of the virion envelope and envelope protein spikes [28].During early infection, the nonstructural proteins are synthesized directly from the two third of genomic RNA as a P1234 polypeptide that is further cleaved to form Nsp1, Nsp2, Nsp3, and Nsp4 nonstructural proteins. Nsp1 protein is involved in the synthesis of minus-strand RNA, in addition to building association of the replication complex with cellular membranes. Nsp2 protein acts as a helicase and proteinase that cleaves the nonstructural polyprotein to form the individual
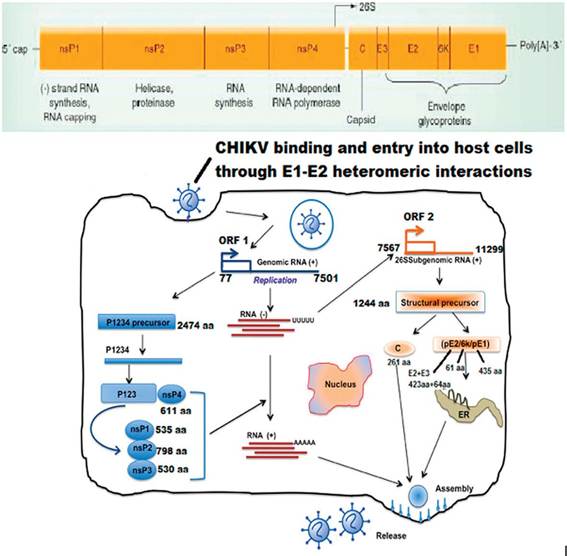
Figure 3.
CHIKVgenome, binding, and entry into host Cellsfollowed by replication.
nonstructural proteins [30]. The function of Nsp3 in viral replication is largely unknown, however, it is probably involved in RNA synthesis [28]. Nsp4 interacts with the N terminal region and other nonstructural proteins and host factors, and acts as the viral RNA polymerase. The sub-genomic mRNA (26S) synthesizes single polypeptide that comprises the structural proteins, such as capsid, E3, E2, 6K and E1. These proteins are then cleaved co-translationally and post translationally to form the functional structural proteins.
These structural proteins have important functions during virus replication and particularly, in the interaction with the host. Such interaction with the host was first presented by the production of antibodies that played important roles in the recovery from infection [31].The region between the nonstructural and structural domains is called the “junction region,” which enhances the transcription of an intracellular subgenomic 26S RNA. There are two other untranslated regions, along with the junction region; one at the 5'-end, which is required for the synthesis of the plus-strand [25], and the other at the 3' end between the stop codon of the E1 gene and the poly (A) tail. This region is mainly involved in translation of viral proteins rather than in replication of the genomic RNA [28, 32].
1.2.3 Replication cycle of CHIKV
The interaction between the envelope proteins of CHIKV and receptors of host cells is required to penetrate into vertebrate cells. The cellular receptors for CHIKV are still unknown; however, in other Alphavirus the laminin receptor, glycosaminoglycans and DC-SIGN (CD209) molecules are involved in viral uptake [33]. The virus is transported into the cell by endocytosis of clathrin-coated vesicles. The activation of E1 protein from the E1-E2 complex is initiated because of the pH reduction of the vesicle, thereby initiating fusion of viral and endosomal membranes, resulting in the release of the nucleocapsid into the cytoplasm. Replication of CHIKV occurs in the cytoplasm. The first event is P1234 precursor polyprotein translation and RNA replication. P1234 polyproteins are directly translated from the viral genome, followed by the initiation of RNA replication through the synthesis of a complete minus-strand RNA, which serves as the template for the synthesis of the viral genome and for the transcription of 26S subgenomic plus-strand RNA from the internal promoter of the junction region. As both processes are interlinked, Nsp4 associates with P123 and other host factors to regulate the synthesis of minus-strand RNA, after cleavage from the P1234 polyprotein.
This switching from genome replication to transcription of sub-genomic 26S positive-strand RNA is also regulated by the nonstructural proteins that were cleaved from the P123 polyproteins [34]. The 26S subgenomic RNA that serves as the mRNA translates the structural protein precursor, and further undergoes co-translational cleavage to become mature (C-E3-E2-6 k-E1). Autocatalytic cleavage of the N-terminal region of structural polyprotein precursor generates the capsid protein, followed by encapsidation of the viral genomic RNA, thereby, resulting in the rapid assembly of nucleocapsid cores in the cytoplasm. In parallel, E2 and E1 are transferred to the plasma membrane after being cleaved from the envelope polyprotein precursor. Finally, the packaging of the virus is performed in the cytoplasm by the assembly of nucleocapsid cores along with glycoproteins, and the virus is released by budding through the cellular membrane to form an enveloped virion [34].1.2.4 Clinical presentations of CHIKV
The most common symptom in chikungunya disease is painful polyarthralgia, mainly bilateral, symmetrical and culminates within few days usually affecting peripheral joints like ankles, toes, fingers, elbows, wrists and knees. The joints exhibit extreme tenderness and swelling with patients frequently reporting incapacitating pain that lasts for weeks or months. Other typical signs for CHIKV infection include fever, headache, retro-orbital pain, chills, weakness, lumbar back pain, joint stiffness, malaise, nausea and a rash that may or may not be accompanied by other signs and symptoms of the disease [35]. The acute illness lasts 3-5 days, with recovery in 5-7 days. The incubation period following the bite of an infected mosquito is short (2-6 days) and ends with a sudden onset of fever reaching as high as 104°F that may last up to 10 days. The fever almost always precedes the rash and joint pain and only very rarely has been reported as biphasic with recurrence noted on the fourth or fifth day of illness.
The rash, appearing primarily on the trunk, face, and limbs of the body is visible on day 2-5 postinfection, and may last up to 10 days. Older patients with an history of rheumatism exhibit more severe symptoms in comparison to younger patients [36].1.2.5 Pathogenesis and diagnosis of CHIKV
Detailed studies on the pathogenesis of the chikungunya fever are rare. It is hypothesized that after inoculation, primary viral multiplication occurs in lymphoid and myeloid cells. The arthropod vectors acquire the virus by sucking blood during this period. The virus, then spreads to the targeted organs and immune system starts functioning at this stage, leading to the activation of both humoral and cellular immunity. This response of the body leads to the development of clinical features of the disease [36].
The probable diagnosis of chikungunya fever can be made on the basis of the presence of the virus in the community, and a clinical trial of fever, rashes and arthralgia, which are suggestive of the illness. The virus produces neutralizing and haemagglutination inhibiting (HI) antibodies, which helps in serological diagnosis. HI test is the simplest diagnostic test; however, it identifies the group rather than specific virus. Confirmation of the illness is done by detection of the antigen or antibody to the analyte in the blood sample of patient [37]. Reverse transcriptase polymerase chain reaction (RT-PCR) is a confirmatory test for the identification of CHIKV IgM capture ELISA is the most sensitive serological assay, and can distinguish the chikungunya from dengue. All virus isolation procedures need to be done under bio safety level 3 (BSL-3) precautions, although such precautions may not be necessary in the countries where CHIKV is endemic.
1.2.6 Transmission cycles of CHIKV
CHIKV is transmitted by mosquitoes belonging to genus Aedes. The mosquitoes considered to be the main vectors for CHIKV are Ae. albopictus and Ae. aegypti [3]. Continuous variations in the geographic distribution of these vectors have been documented in several studies. Ae. aegypti was considered to be the primary vector of CHIKV in most parts of the globe [38], whereas Ae. albopictus (common name Asian tiger mosquito) was considered to be the secondary vector and was restricted to Asia [38]. However, the recent reemergence of CHIKV in many parts of the world has been mainly associated with Ae. albopictus vector [19, 38]. Furthermore, reports indicate Ae. albopictus to replicate and transmit the old African genotype of CHIKV as well as the recent Indian Ocean strain of CHIKV better than those of Ae. aegypti and other Aedes species [39]. CHIKV is endemic in tropical regions of Africa and Asia, where the mechanisms of CHIKV transmission and maintenance appears to be very complex and vary significantly depending on the particular region where virus activity is detected.
1.2.7 CHIKVin African mosquito vectors
In Africa, CHIKV is believed to be maintained in a sylvatic as well as with urban/ rural cycle involving wild nonhuman primates and forest-dwelling Aedes mosquitoes (Figure 4). Several field studies conducted in Senegal, Nigeria, Uganda, Tanzania, Cote d’Ivoire, Central African Republic and South Africa concluded that the main sylvatic vectors of CHIKV are probably Ae. furcifer, Ae. taylori, Ae. africanus, Ae. luteocephalus and Ae. Neoafricanus [32]. Based on the isolation frequencies, it appears that Ae. furcifer and Ae. taylori are more important in southern and western Africa, while Ae. africanus is the chief vector in central regions [35]. Laboratory studies have confirmed vector competence of Aedine sylvatic mosquitoes in Africa. In South Africa, the oral infectious dose of Ae. furcifer was 50% (OID50)—the titer of the virus in the blood meal sufficient to infect 50% of mosquitoes was found to be less than 6.2 log/ml resulting in a transmission rate of 25-32%. This is sufficient to sustain CHIKV transmission from velvet monkeys and baboons, which develop viremia up to 7-8 log/ml [41].
1.2.8 CHIKV circulation in Asian mosquito vectors
In contrast to Africa, only urban/rural CHIKV transmission cycle has been described in Asia (Figure 4). Ae. aegypti is the main vector of CHIKV and Ae. albopictus is believed to play a secondary role in several outbreaks. CHIKV epidemics in humans seem to be disconnected from zoonotic transmission, however; the recent study of seroprevalence to CHIKV infection among wild monkeys in the Philippines showed presence of anti-CHIKV IgG in 59.3% of animals tested, suggesting existence of a possible sylvatic transmission cycles [42]. Currently, it is believed that persistence of CHIKV in Asia results from viral migration back and forth among different locations sustained by the human-Ae. aegypti cycle [43].
The vectors responsible for viral transmission during these epidemics have not been definitely characterized. Both Ae. aegypti and Ae. albopictus are present in India and their epidemiologic significances for CHIKV transmission probably vary
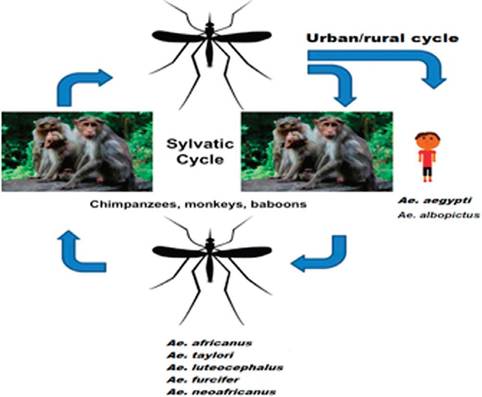
Figure 4.
CHIKV transmission cycle in Asia and Africa (modified from Thiboutot et al. [29, 40]).
dependent on the geographic location. Another intriguing feature of the 2006-2008 CHIKV epidemic in India, beside the magnitude, is the fact that this epidemic was caused by virus of the ECSA genotype. All previous outbreaks were caused by Asian genotype of CHIKV It was proposed that this shift in viral genotype was the major factor in the re-emergence of Chikungunya in an unprecedented outbreak in India after a gap of 32 years [44].
1.3