Dengue: overview
1.3.1 History and geographic distribution of DENV
DENV is found in tropical and subtropical areas throughout the world, with prevalence in both urban and suburban areas. DENV is endemic in more than one-hundred countries with more than two-and-a-half billion people and around 40% of the world’s population living in areas at risk for infection.
The World Health Organization estimates that there are between fifty and one-hundred million DENV infections each year, causing hospitalization of five-hundred thousand people, and a death rate of two-and-a-half percent [45]. The earliest report of disease with dengue-like symptoms dates back to a Chinese encyclopedia of disease symptoms and remedies that was published from 265 to 420 A. D during the Chin Dynasty [46]. It is speculated that DENV was the etiological agent during disease outbreaks in the French West Indies in 1635, in Panama in 1699, and the Philadelphia epidemic of 1780 [47]. Reported cases of dengue disease were seen in 1779 and 1780 in Africa, Asia, and North America [48]. The first verified dengue epidemic occurred from 1953 to 1954 in the Philippines followed by a quick global spread of epidemics of DF/DHF. In the 1980s and 1990s, DENV continued to expand, and reached areas with mosquito vectors [49].The very first report of the existence of dengue fever in India was in 1946 from US soldiers in Kolkata [50]. Since then, there was no significant dengue activity reported anywhere in the country for the next 18 years. In 1963-1964, an epidemic of dengue fever was reported from the Eastern Coast of India, further spreading northwards and reached Delhi in 1967 and Kanpur in 1968. Simultaneously, the DENV epidemic also engulfed the southern part of the country and gradually the whole country was affected by wide spread epidemics followed by endemic/ hyperendemic prevalence of all the four serotypes of DENV However, most dengue outbreaks in India were simple dengue fever with very rare cases of DHF/DSS epidemics.
The first major wide spread epidemics of DHF/DSS occurred in 1996 in India, involving areas around Delhi and Lucknow, further spreading across the country [51, 52]. Since then, the epidemiology of DENV and its prevalent serotypes has been frequently changing in India.1.3.2 Dengue virus (DENV)
Dengue virus (DENV) is a member of enveloped, positive-strand RNA viruses of the flaviviridae family. The flaviviridae family also includes West Nile virus, yellow fever virus, Japanese encephalitis virus, hepatitis C virus, and tick-borne encephalitis virus. Flaviviruses are transmitted to humans by arthropod vectors such as mosquitoes or ticks [49].
1.3.3 Dengue viral structure
There are four phylogenetically and genetically distinct, but antigenically related serotypes classified as DENV-1, DENV-2, DENV-3 and DENV-4. The dengue virion is a spherical particle, existing as either a 50 nm diameter immature particle or a mature 60 nm diameter particle with a lipopolysaccharide envelope. DENV genome is about 11 kb with a single ORF encoding three structural proteins: capsid (C), membrane (M), and envelope (E) and seven viral encoded nonstructural proteins: NS1, NS2A, NS2B, NS3, NS4A, NS4B and NS5 (Halstead, 2008). DENV RNA contains a type I cap structure (m7GpppAmpN2), located at its 5'-end, and lacks the poly (A) tail at its 3' end. The DENV genome is surrounded with C proteins and forms the inner core. The structural proteins E and M are surface proteins on the virion envelope and the conformations of these proteins are used to distinguish between immature and mature virus. The immature virus is referred to as “spiky” as M proteins bound to a precursor membrane protein (pr) form heterodimers with E proteins that appear as “spikes” on the viral surfaces. In mature virions, the soluble pr is cleaved from M protein by furin, anchoring the M proteins and causing the pr protein to be absent in the mature viral membrane [53].
1.3.4 Dengue viral replication
DENV enters a variety of cells including macrophages, monocytes, and dendritic cells though cell endocytic vesicles.
The first step in DENV infection is binding to the cellular receptors on the surface of the target cell like ubiquitous receptor (DC-SIGN). This interaction leads to the internalization of the virion via receptor-mediated endo- cytosis, resulting in the fusion of the virus with the endosomal membrane because of acidification, and finally releasing the viral genome into the cytoplasm. DENV genome is associated with rough ER (site of its translation), because the viral RNA can act as mRNA. Viral replication occurs in two steps: first, the positive-polarity RNA is copied to RNA of negative polarity that serves as a template for the synthesis of multiple strands of RNAs of positive polarity (amplification process); the positivepolarity RNA can then translate into proteins, for further RNA synthesis of negative polarity, or can become associated with structural proteins C, E, and M to form the viral progeny. Second, the immature virus particles travel to the Golgi apparatus in vesicles, where they undergo glycosylation, and are finally transported through secretory vesicles outside the cell (Figure 5). During the latter process, the furin cleaves prM in M to generate mature virions, which is the final step of viral morphogenesis [55]. The three main elements that are necessary for DENV replication are: cis-acting elements, trans-acting factors, and viral induced membranes.1.3.4.1 Cis-acting elements
The cis-acting elements are mainly located at both ends of DENV genome in the 5'- and 3'-UTR. The cyclization sequences, as well as the upstream UAG region located at both ends of DENV genome and the downstream AUG region induce circularization of DENV genome. For an efficient negative-strand RNA synthesis, the secondary structure of the stem loop at the 3'-end (3'-SL), along with the secondary structure of SL structures within the 5'-UTR are essential. The initiation of viral replication occurs with the binding of NS5 (RNA dependent RNA-polymerase) to the 5'-UTR.
Trans-acting factors are of two types: viral trans-acting factors and cellular transacting factors.
Viral trans-acting factors: NS3 and NS5 (multifunctional and multidomain proteins, respectively) are the only proteins encoded by DENV possessing catalytic activities. NS5 has two main activities: RNA-dependent RNA-polymerase and methyltransferase. NS3 has protease, helicase, and nucleoside triphosphatase activities. NS3 functions by regulating its association with other viral proteins. NS1 and the small nonstructural proteins are required for anchoring the viral replication complex to the membranes of the endoplasmic reticulum.
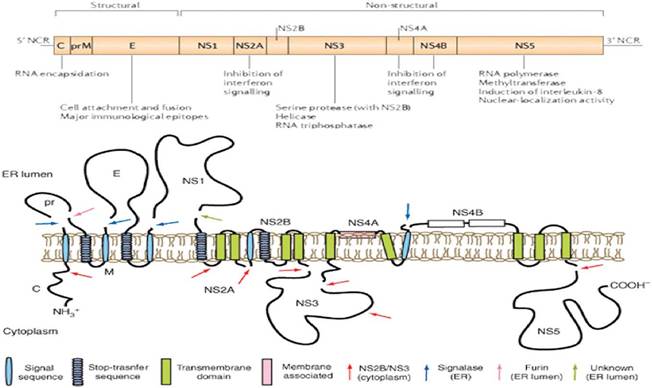
Figure 5.
Picture demonstrating: gene organization in DENV RNA genome (top), membrane topology and proteolytic cleavage sites of the transcribed polyprotein (bottom). Arrows denote the cellular and viral proteases, which process the immature polyprotein into ten separate proteins (modified from Perera and Kuhn [54]).
Cellular trans-acting factors: several cellular proteins, such as elongation factor 1a (EF1a), polypyrimidine tract binding protein (PTB), LA, calreticulin, PDI, and the heterogenous nuclear factors A1, A2/B1 and Q, have been found to bind to the 5'- or 3'-UTR of DENV During DENV infection, PTB and La proteins translocate from the nucleus to the cytoplasm and act as the positive and negative regulators of viral replication, respectively. The YB-1 protein might participate in the switching from viral translation to replication or might have a role as an antiviral factor [54, 55].
1.3.4.2 Viral induced membranes (replication complex)
For the formation of the replication complex, proliferation and the generation of invaginations of the ER membranes are required initially, which are probably induced by NS4A and NS3 in conjunction with cellular and other viral proteins. Invaginations are mainly considered as the site for viral replication. The DENV RNA is exported to the convoluted membranes that might potentially store proteins and lipids required for DENV replication.
Viral morphogenesis is initiated by the association of the RNA and the C protein generating nucleocapsids. The C protein accumulates around the lipid droplets in the ER. Accumulation of immature viral particles occurs in the lumen of dilated ER cisternae, which are then transported to the cis-Golgi for maturation (Figure 6) [56].1.3.5. Classification and symptoms of dengue
Cases of symptomatic dengue have historically been classified by severity according to WHO guidelines first published in 1975, which differentiate between cases of dengue fever (DF), dengue hemorrhagic fever (DHF) and dengue shock syndrome [3] (Figure 7). However, dengue epidemiology has changed considerably since these guidelines were first published, leading to difficulties with the use of this classification system in a clinical setting. Examples of severe dengue that do not follow WHO criteria of DHF stratification are dengue with hemorrhage but without evidence of plasma leakage; dengue with shock syndrome without fulfilling all four DHF criteria; and severe dengue accompanied with organ dysfunction and a low level of plasma
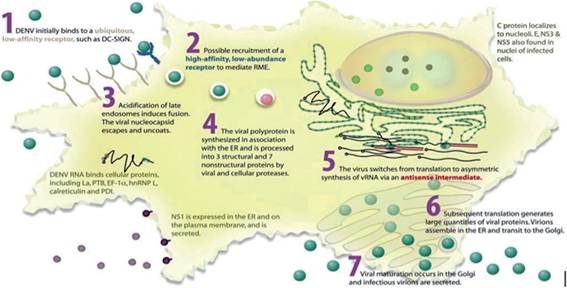
Figure 6.
Dengue replication cycle. Dengue enters a susceptible cell through receptor-mediated endocytosis. In endosomic vesicles, dengue virions are uncoated and release the genome into the ER. Viral RNA is translated into a polypeptide and processed to form viral proteins. Replication and viral assembly occurs in the ER, and the virions travel to the Golgifor modification and is exported via exocytic vesicles (adapted from Clyde et al. [56]).
leakage. In view of the above facts, recently the WHO Tropical Disease Research (TDR), 2009 [57] proposed a new classification of dengue, i.e., dengue (D), dengue with warning signs (DW) and severe dengue (SD) in order to re-evaluate the current classification for better management of high case fatalities. However, the previous
Dengue fever
Acute febrile illness with two or more of the following:
• headache
• retro-orbital pain
• myalgia
• arthralgia
• rash
• haemorrhagic manifestations
• leukopenia
and either supportive serology or proximity to laboratory confirmed cases.
Dengue haemorrhagic fever
The following indications must all be present:
• fever or a history of fever
• haemorrhagic tendencies evidenced by one or more of the following:
- a positive tourniquet test
- petechiae7 ecchymoses or purpura
bleeding from the mucosa, gastrointestinal tract, injection sites or other locations
- haematemesis or melaena
• thrombocytopenia (100,000 cells per mm3 or less)
• evidence of plasma leakage due to increased vascular permeability, manifested by at least one of the following:
a rise in the haematocrit equal to or greater than 20% above average for age, sex and population
- a drop in the haematocrit following volume-replacement treatment equal to or greater than 20% of baseline
- signs of plasma leakage such as pleural effusion, ascites and hypoproteinaemia
Dengue shock syndrome
All four of the above DHF criteria must be present, plus evidence of circulatory failure manifested by either of the following:
• rapid, weak pulse and narrow pulse pressure (intra serotypic recombination, mutation and ecological changes, thereby resulting in generation of new genotypes, which are more virulent, resistant and can cause massive outbreaks affecting large number of people. Introduction of such new viral genotypes inevitably leads to the question whether particular genotypes of DENV are associated with higher virulence or severe disease. To date, several diseases have often been associated with several DENV genotypes originating in Southeast Asia. The lack of a suitable animal model for the dengue disease poses challenges in confirming such hypothesis [64].
1.4