Diseases Caused by Allogeneic Incompatibilities (Horses and Ruminants)
M. Julia B. Felippe • Jill R. Johnson
The surfaces of cells are covered with molecular structures, the presence or absence of which is determined by genes and cell function. Some of these molecules have limited heterogeneity within a species, and all members of the species share identical forms (monomorphic).
In other cases, a spectrum of minor structural variations or polymorphisms of a particular molecule occur within the population; therefore some members of the species will have one form of the molecule, whereas others will have another. If the structural variations are such that the immune system of one individual can recognize the differences in the molecules from another individual, these are known as alloantigens. Antibodies produced against these antigens are alloantibodies. Examples of alloantigens include certain molecules of RBCs, platelets, and WBCs.In nature, because there are few situations in which tissues are exchanged, there are relatively few naturally occurring diseases that involve allogeneic incompatibilities. Neonatal isoerythrolysis (NI), neonatal alloimmune thrombocytopenia (NAIT), and alloimmune neonatal neutropenia (ANN) are three examples of such naturally occurring diseases. Blood transfusion reactions and organ transplant rejection are examples of iatrogenic diseases associated with allogeneic incompatibilities.
The same alloantibodies that mediate disease are also useful in vitro as reagents for the detection of the presence of alloantigens and serve as the basis for blood typing, for instance.
Blood Typing and DNA Profiling
RBC alloantigens, major histocompatibility complex molecules, and electrophoretic markers. Polymorphisms of the DNA have emerged as useful genetic markers and essentially replaced other genetic tests for parentage verification.
Red Blood Cell Antigens
The blood group genes produce surface molecules that contain antigenic sites known as factors.
More than 30 different factors have been identified. Each factor is associated with only one system, although the same factor may be associated with more than one allele within the system. The factors are named with an uppercase letter to denote the system and a lowercase letter to designate the factor within that system. Groups of factors that are produced by a single allele are called phenogroups.The presence of RBC antigens is detected primarily by either agglutination or complement-mediated lysis of test cells using antibodies directed against specific RBC alloantigens. In some systems, antigens are detected with antiglobulin tests.
Seven independent RBC groups or systems have been internationally defined in horses under the auspices of the International Society for Animal Genetics1-4 (Tables 53.8 and 53.9). These systems are named A, C, D, K, P, Q, and U. Additional systems are recognized by individual laboratories. Each system corresponds to a particular gene for which two or more alleles exist. Some blood groups are relatively simple and contain only two alleles, whereas other systems may have more than two dozen different alleles.
Eleven blood groups have been identified in cattle5 (Tables 53.8 and 53.10). In cattle, groups B and J have the greatest clinical relevance. The B group is a complex system with more than 60 different antigens.6 This complexity has been used to advantage for individual animal identification and parentage studies in the past, but it makes it difficult to closely match donor and recipient blood for transfusions. The J antigen, a lipid found in body fluid, is adsorbed to erythrocytes; thus it is not a true erythrocyte antigen. Newborn calves do not have this antigen but usually acquire it during the first 6 months of life. After acquiring it, some groups of cattle have high amounts of J antigen both in the serum and adsorbed to erythrocytes, whereas other groups have small amounts found only on erythrocytes.
These latter groups, often referred to as J negative, actually may have anti-J antibodies and develop transfusion reactions when transfused with blood from J-positive donors.1Seven blood groups have been identified in sheep (see Table 53.8). The B system is analogous to the B system in cattle and is extremely polymorphic, with more than 52 different alleles. The R system is similar to the J system in cattle in that the antigens are soluble and are passively adsorbed to erythrocytes.5 The M-L system is involved in the genetically determined RBC potassium polymorphism found in sheep.7
Blood groups of goats appear to be similar to those of sheep, and many of the reagents used for typing sheep blood have been used to type goats' blood. Reagents used to detect antigens
■ TABLE 53.8
Domestic Animal Blood Groups
| Species | Blood Group Systems |
| Equine | A, C, D, K, P, Q, U |
| Bovine | A, B, C, F, J,a L, M, R', S, T, Z |
| Ovine | A, B, C, D, M, R,a X |
| Caprine | A, B, C, M, Ja |
aSoluble antigens.
From Suzuki Y: Personal communication, Stormont Laboratories, Inc., Woodland, Calif., 1990.
■ TABLE 53.9
Blood Group Systems, Factors, and Alleles of the Horse Recognized by the International Society for Animal Genetics
| System | Factors | Recognized AUeles |
| A | a,a b,b c, d, e, f, g | Aa, Aadf, Aadg, Aabdf, Aabdg, Ab, Abc, Abce, Abe, Ac, Ace, Ae, A- |
| C | a | Ca, C- |
| D | a, b,b c,b d, e, f, g,b h, i, | Dadl, Dadlnr, Dadlr, Dbcmq, Dcefgmq, Dcegimnq, Dcfgkm, Dcfmqr, Dcgm, Dcdmp, |
| k, l, m, n, o, p, q, r | Dcgmq, Dcgmqr, Ddeklqr, DeIno, Ddeloq, Ddelq, Ddfkir, Ddghmp, Ddghmq, Ddghmqr, Ddki, Ddlnq, Ddlnqr, Ddlqr, Ddno, Dq,(D-) | |
| K | a | Ka, K- |
| P | a,b b, c, d | Pa, Pac Pacd, Pad, Pb, Pbd, Pd, P- |
| Q | a,a b, cb | Qa, Qab, Qabc, Qac, Qb, Qbc, Qc, Q- |
| U | ab | Ua, U- |
aMost common factors involved in neonatal isoerythrolysis (NI).
bPreviously reported to cause NI in at least one case.From Cothran G: Personal communication, University of Kentucky, Lexington, Ky., 1996.
■ TABLE 53.10
■ TABLE 53.11
Blood Group Factors of Cattle Recognized by the International Society for Animal Genetics
| System | Factors |
| AAa | A1, A2, H, Z, a |
| Fa | F, V |
| J | J, j |
| L | L, l |
| M | M, m |
| Z | Z, z |
| R" | R', S' |
| Bb | B, G1, G2, G3, l1, l2, K, O1, O3, OX, Pl, P2, Q, T, Y2, A', B', D', E1', E2', E3', G', I1' (I'), I2', Jl', J2', K', O', P1', P2', Q', Y', A", B", G", I" |
| C | C1, C2, R1, R2, W, X, X2, L', E1, C', C", X', Fl, F6, F10, Fl5 |
| S T | S1, S2, U, H', U', S", U" |
aCases of neonatal isoerythrolysis associated with production of anti-red blood cell antibodies against factors in response to anaplasmosis and Babesia vaccines. bAdditional factors are recognized by individual laboratories.
Examples of Polymorphic Proteins Used in Blood Typing
| Protein | System | Source |
| Alpha1 B glycoprotein | AlB | Serum |
| Albumin | ALB | Serum |
| Transferrin | TF | Serum |
| Carboxylesterase | ES | Serum |
| Vitamin D-binding protein | GC | Serum |
| Protease inhibitor | PI | Serum |
| Peptidase A | PEPA | Serum |
| Plasminogen | PLG | Serum |
| Glucose phosphate isomerase | GPI | RBCs |
| 6-Phosphogluconate dehydrogenase | PGD | RBCs |
| Phosphoglucomutase | PGM | RBCs |
| Catalase | CAT | RBCs |
| Carbonic anhydrase | CA | RBCs |
| Acid phosphatase | AP | RBCs |
| Hemoglobin A | HBA | RBCs |
| NADH diaphorase | DIA | RBCs |
NADH, Reduced nicotinamide adenine dinucleotide; RBC, red blood cell.
of the J system of cattle have been used to detect differences in a similar system in goats.8
The biological function of RBC antigens is not known for any system in any species with the exception of the M-L blood group system in sheep, which is involved in active potassium transport across RBC membranes.
Electrophoretic Markers
Many plasma and intracellular proteins have polymorphic forms. These variants have subtle biochemical differences, which for the most part do not alter the major characteristics of the molecules but can be detected using electrophoretic methods. These are not technically alloantigens because the differences are not detected immunologically. The various forms of the molecules are detected based on their different rates of migration in electrophoresis. The specific forms of any particular protein that an individual possesses are determined genetically, and those used in blood typing are usually expressed codominantly. Currently, no diseases are clearly associated with the presence or absence of particular alleles of polymorphic proteins; however, they are useful genetic markers for identification and parentage studies commonly included in the blood type (Table 53.11).3,8-10 Electrophoretic markers were used extensively in horses because RBC antigens did not provide the same degree of discrimination among individuals in this species as they do, for example, in cattle. The current application is limited to legacy parentage verification when parental DNA is not available for testing.
Major Histocompatibility Complex
Three systems of equine alloantigens tested using lymphocytes have been described: ELA, ELY-1, and ELY-2.11-15 ELA is the major histocompatibility complex (MHC) of the horse. ELA is a complex system that includes several genes, each of which is polymorphic (e.g., has many different alleles). Because the MHC regulates the interactions of many cells in the immune response, a potential for the association of disease susceptibility or resistance with ELA exists. An association between a particular ELA allele and equine sarcoid has been described.16,17 ELY-1 and ELY-2 have limited polymorphism and appear to be two allele systems.
The MHC of cattle is called BoLA, for bovine lymphocyte antigen.
In sheep the MHC is called OLA, for ovine lymphocyte antigen, and is also known as SH-LA. In goats the comparable system is GLA.DNA Polymorphisms
A wide variety of approaches has been developed for detecting genetic variation using DNA. Certainly, DNA sequencing is the most sensitive approach, but other approaches have been used to detect DNA variants. Genetic variation of DNA can be detected among individuals based on differences in length for a section of DNA or even for differences in base composition at specific sites. DNA length variants are usually referred to as minisatellite or microsatellite markers. Minisatellite DNA markers are based on sequences, often 15 to 100 bases long, that may be repeated hundreds of times on a chromosome. The number of repetitions can vary dramatically among individuals and collectively is referred to as DNA fingerprinting. Although they are powerful, these tests are costly and time consuming. More recently, another type of DNA length variant has been discovered. Microsatellite DNA markers are short repetitions of nucleotides, usually ranging from 1 to 9 bases long, which may be repeated 15 to 40 times. As with minisatellites, the number of repetitions may vary among chromosomes. More than 100,000 of these markers are thought to exist in each species and to contribute to the development of genetic maps in diverse species. These markers currently form the basis for most parentage analysis.
Variation recognizable as a change in a single base of DNA is more subtle but can be detected through a variety of techniques. Single-nucleotide polymorphisms (SNPs) are variations in single nucleotides at specific positions in the genome, such as coding or noncoding sequences of genes, or between genes (intergenic). These variations may not change (synonymous SNPs) the protein sequence; however, they may change (nonsynonymous SNPs) the protein sequence or alter its expression. SNPs are used to determine haplotypes (linkage of SNPs) in a population, as well as in association studies for individual genetic background and susceptibility to certain diseases, disease severity, and effective treatments.19,20 Examples described for horses include susceptibility to insect bite hypersensitivity, uveitis, R. equi pneumonia, and papillomavirus- induced sarcoids in horses, as well as variations in the immune system.21-28 Examples described for ruminants include susceptibility to mastitis, ketosis, tuberculosis, paratuberculosis, as well as variations in the immune system.29-35
DNA sequence variation is often detectable using restriction enzymes. Restriction enzymes are isolated from bacteria and will cleave DNA when the sequence is specific for the enzyme. For example, the method to detect the gene defect causing hyperkalemic periodic paralysis (HYPP) in horses is based on polymerase chain reaction (PCR) amplification of the gene and then detection of sequence differences that occur between the normal and mutant gene using restriction enzyme digestion of the DNA. Other approaches to identifying DNA sequence differences include single-strand conformational polymorphism (SSCP), allele-specific PCR amplification, and ligase chain reaction. DNA-based testing has essentially replaced blood typing for purposes of parentage analysis.
Blood Typing and DNA Genotyping Applications chromosomes are expressed. This often allows determination of genotype (the actual genes present) from the phenotype (the antigens actually detected). Some systems have null alleles with no detectable products that would allow inference of genotype (e.g., C, K, and U blood group systems in horses). An individual can have only two of the possible different alleles for any given gene (one on each chromosome of a pair).
Blood typing based on RBC antigens and electrophoretic markers has been used for animal identification and pedigree analysis, but currently RBC typing is used primarily for prediction of potential for NI and crossmatching for blood transfusion.
The use of genetic markers for parentage verification is based on exclusion (i.e., markers are tested until a discrepancy is found). If no discrepancy is observed after a statistically acceptable amount of testing, the probability of a match is determined.
Because genetic testing is a composite mosaic of a large number of different, largely independent genetic markers, the odds of two individuals sharing exactly the same pattern of markers, whether RBC antigens, electrophoretic markers, or DNA polymorphisms, are remote. Thus once an animal's genetic type is on record, it can be a powerful identifier of an individual animal. In this sense it is an unalterable form of identification that is present from birth to death.
These markers can be used to support or refute parentage claims because the presence or absence of markers is genetically determined and because the markers contributed by both parents are expressed in a codominant fashion (unless “blank” alleles exist). Blood typing and DNA profiling cannot prove that one horse is the parent of another; however, in certain instances it can exclude with certainty a stated parent as the true parent. Two types of parentage exclusions can occur: type I and type II36 (Fig. 53.7). With type I, the foal possesses a dominant or codominant marker that is not present in either parent; this result indicates that at least one of the stated parents is incorrect. If one parent (usually the dam) is known with certainty based on circumstances other than genetic type, the error lies with the other stated parent. With type II exclusions, the foal does not possess either of the dominant or codominant markers of a stated parent. Because a foal must inherit one of the two markers expressed by each parent, exclusion can be made if neither of the two markers present on the stated parent is detected in the foal. In parentage cases, each genetic system is evaluated for type I and type II exclusions. A parent can be excluded based on an inconsistency in one system. For this reason, the more systems that are used, the better genetic typing becomes as a tool for solution of parentage disputes. The calculated effectiveness of blood typing for detecting incorrect paternity using 20 internationally defined systems in horses is as high as 96%.3,9,37 With DNA genotyping, effectiveness is as high as 99.998%.38-40
Laboratories that provide DNA genotyping in North America are listed in Box 53.1.
Neonatal Isoerythrolysis
■ Equine
Definition and Etiology
Neonatal isoerythrolysis (NI) is a condition characterized by the destruction of RBCs in the circulation of a foal by alloantibodies of maternal origin absorbed from colostrum.2,41,42
Production of alloantibodies can be stimulated in mares in several ways, including transfusion, exposure to blood from the foal during parturition, or as a result of placental pathology during gestation. Incompatibilities of at least some blood groups between the dam and foal are the rule rather than the exception, and yet the incidence of sensitization of the dam and occurrence of NI is relatively low. Most blood groups are not strongly antigenic under the conditions of exposure via previous
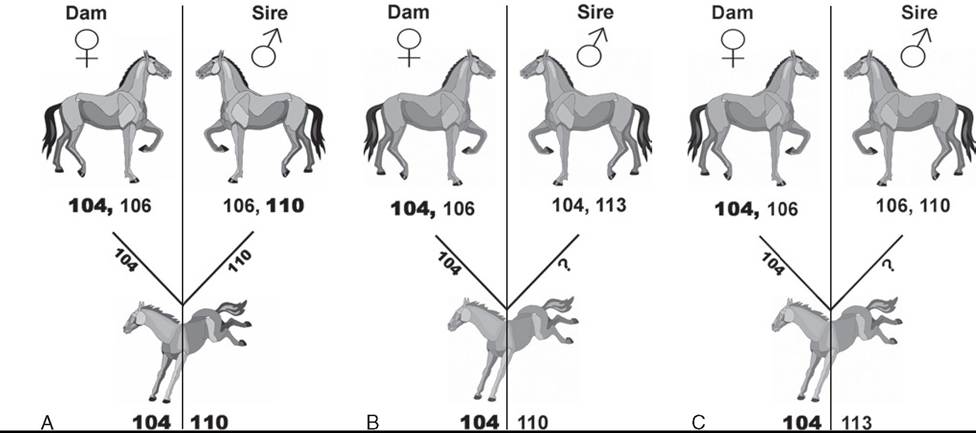
FIG. 53.7 A, The sire of this foal cannot be excluded as the sire of the foal based on the results for the DNA marker VHL20. B, The sire of this foal is excluded by a type I exclusion. The foal possesses an allele for the DNA marker VHL20 not present on either parent. Assuming that the dam of the foal is known to be correct and therefore would have contributed the 104 allele to the foal, the sire must be incorrectly stated. C, The sire of this foal is excluded by both a type I and a type II exclusion. The foal possesses an allele not present on either parent and also does not possess either allele expressed by the putative sire.
parturition or placental leakage. Several blood group factors, however, are particularly immunogenic, and antibodies against these antigens have been reported to cause nearly all cases of NI. These include factor Aa in the A system and factor Qa in the Q system. In mules, a unique donkey RBC antigen called donkey factor has been associated with NI.43,44 Antibodies to these antigens develop after the exposure of the mare to RBCs and are not naturally occurring. In humans, the absence of certain RBC antigens results in the production of natural antibodies against that antigen, presumably because of exposure to cross-reacting antigens in the diet. This does not occur in horses, with a few exceptions, and has not been associated with NI. Ca-negative (Ca-) horses frequently make anti-Ca antibody of low titer without known RBC exposure. However, anti-Ca antibody is generally not associated with NI, and mares with anti-Ca antibody appear less likely to develop certain antibodies responsible for NI.
For NI to occur, several things must happen (Figs. 53.8 and 53.9). First, the dam must be negative for the antigen in question. Mares that are Aa-, Qa-, or both are considered at risk because most cases of NI in horses are associated with either anti-Aa or anti-Qa antibodies. All horses appear to lack donkey factor; thus all mule pregnancies are considered at risk. Second, the mare must become sensitized and produce antibody to the offending antigen. Sensitization can be caused by exposure during a previous pregnancy, blood transfusion, or transplacental contamination with fetal RBCs earlier in the current pregnancy (rare). Third, the foal from the current pregnancy must have inherited from its sire the antigens to which the mare has been sensitized. When these conditions are met, there is a significant potential that maternal antibody directed against the foal's RBC antigens will appear in the colostrum and, if subsequently ingested and absorbed by the foal, could cause loss of RBCs. The higher the anti-RBC titer is at foaling, the higher the risk. The highest titers are likely to be produced in a previously sensitized mare that is reexposed to the same RBC antigens shortly before parturition.
■ Clinical Signs and Differential Diagnoses Foals are born healthy and usually begin to develop signs of NI at 24 to 36 hours of age, after suckling. Progressive lethargy and weakness are early signs. In acute cases, mucous membranes show initial pallor that is followed by icterus. In severe cases, hemoglobinemia and hemoglobinuria may be pronounced. In peracute cases, death may precede the development of icterus. Breathing becomes rapid and shallow, followed by labored breathing as the disease progresses. The foals may yawn repeatedly. Heart rate is elevated. Seizure-like activity may occur as the anemia becomes more severe.
■ Clinical Pathology Affected foals are anemic. All indicators of RBC concentration (packed cell volume [PCV] hemoglobin, RBC count) show significant decreases. PCV values often decline into the teens, and values as low as 5% have been observed. Hemoglobinemia and hemoglobinuria may be present. Bilirubin (mainly unconjugated) levels will be increased as a result of accelerated RBC destruction. Total bilirubin levels may be close to 20 mg/dL in severe cases. Affected foals, especially mule foals, may also be thrombocytopenic.4
Demonstration of significant amounts of alloantibodies in the colostrum (or serum from the mare) that are directed against the RBC antigens expressed by the foal provides a definitive diagnosis of NI. These antibodies are most commonly demonstrated by lytic and agglutinating tests.45,46 Lytic tests are believed to be more reliable indicators of the presence of offending antibody.45 The presence of antibodies attached to the foal's RBCs can also be demonstrated with a direct antiglobulin test (Coombs test). The presence of antibodies in the mare's serum that attach to RBCs can be demonstrated with an indirect antiglobulin test.2
■ Pathophysiology In mares sensitized to RBC antigens, most commonly Aa or Qa, alloantibodies are concentrated in the colostrum late in gestation. These antibodies are passed to the foal through passive transfer. If the foal's RBCs carry the antigen that the antibody recognizes, the cells become antibody coated. Subsequently, they are removed prematurely by the mononuclear phagocyte system (reticuloendothelial system) or lysed intravascularly by complement. A distinction has been made between antibodies that are lysins as opposed
■ BOX 53.1
Laboratories Providing DNA Genotyping Services in North America
Animal Genetic Testing and Research Laboratory
108 Gluck Equine Research Center University of Kentucky
Lexington, KY 40546 (859) 218-1212 www.ca.uky.edu/gluck/AGTRL.asp (Horses)
Animal Genetics, Inc. 1336 Timberline Road
Tallahassee, FL 32312-1766 (866) 922-6436 www.horsetesting.com
(Horses)
Maxxam Analytics Animal DNA Laboratory
335 Laird Road, Unit 2
Guelph, Ontario
N1H 6J3 Canada
(519) 836-2400 http://maxxam.ca/services/dna-testing/animal-dna-testing (Horses and cattle)
Zoetis Genetics
333 Portage Road, Building 300 Kalamazoo, MI 49007-9970 (269) 833-4744 www.zoetis.com/products-services/genetics (Cattle)
Igenity
4665 Innovation Drive, Suite 120
Lincoln, NE 68521 (402) 435-0665 www.igenity.com (Cattle)
Genetic Visions, Inc. 3220 Deming Way #160 Middleton, WI 33562
(608) 662-9170 www.geneticvisions.net (Cattle)
Veterinary Genetics Laboratory University of California, Davis PO Box 1102
Davis, CA 95617-1102 (530) 752-2211 www.vgl.ucdavis.edu (Multiple species)
Molecular Cytogenetics and Genomics Laboratory Department of Veterinary Integrative Biosciences Texas A&M University
Room 314B, Bldg. 1197 (VRB) College Station, TX 77843
(979) 862-2879 or (979) 458-0520 http://vetmed.tamu.edu/vibs http://vetmed.tamu.edu/labs/cytogenics-genomics (Multiple species)
SRC Genserve Laboratories 125-15 Innovation Blvd.
Saskatoon, Saskatchewan
S7N 2X8 Canada
(306) 933-7700 www.src.sk.ca
(Multiple species)
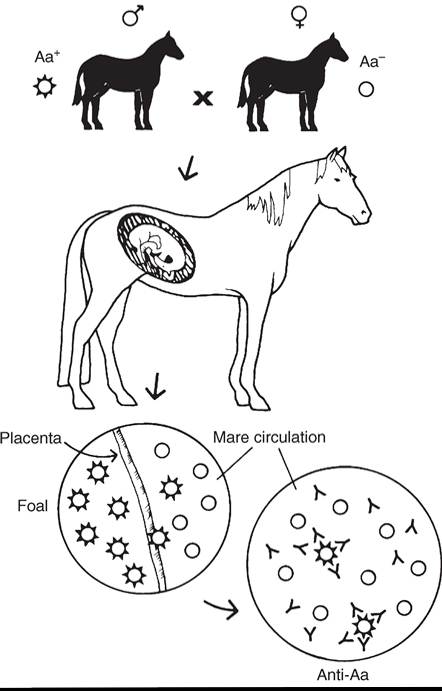
FIG. 53.8 Entrance of red blood cells of paternal antigen type into maternal circulation stimulates the production of alloantibody and, consequently, the presence of alloantibodies in the mare’s serum and in the colostrum.
to agglutinins; however, this distinction is based on in vitro testing and may be somewhat artificial. Under appropriate laboratory conditions, offending antibodies may exhibit both abilities.43 However, when using certain conventional agglutination tests, some alloantibodies capable of producing NI may go undetected.10
■ Epidemiology The percentages of mares at risk for sensitization against the common offending antigens (Aa and Qa) vary among breeds, depending on the frequency in the population of each gene involved (Table 53.12). In Thoroughbreds the prevalence is about 1%, and in Standardbreds the incidence is about 2%.42 The prevalence in mules (donkey sire, horse dam) has been reported to be as high as 10%.2 Increased numbers of mares at risk in a breed do not necessarily translate to higher numbers of NI cases. A corresponding number of stallions lack the factors in question and are therefore unable to sire foals with the offending antigen.42 Virtually all mule
44
pregnancies are incompatible with regard to donkey factor.44
Although the disease is more common in multiparous mares, NI can occur with the first pregnancy.
■ Necropsy Findings Liver failure, kernicterus, and complications related to bacterial sepsis are the most important causes of death in foals dying from NI.47 Pale tissues with or without icterus and splenomegaly are characteristic necropsy findings. Lesions associated with RBC destruction and anemia,
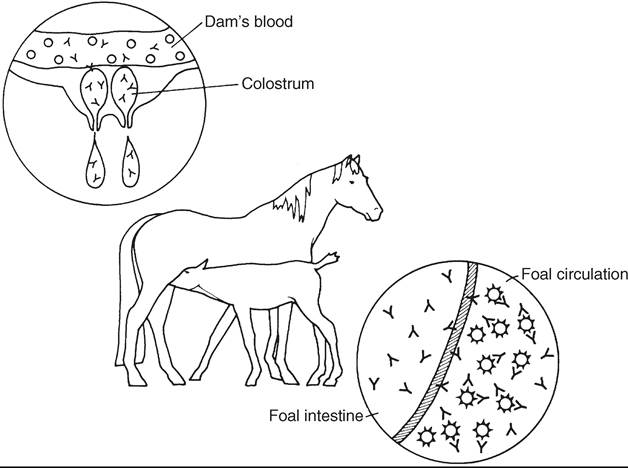
FIG. 53.9 Alloantibody in the mare’s serum is concentrated in colostrum at the end of gestation. Through passive transfer, the foal absorbs immunoglobulins, including alloantibodies. The absorbed antibodies attach to the foal’s red blood cells and cause either premature removal from circulation or intravascular lysis.
■ TABLE 53.12
Estimate of the Percentage of Mares in Selected Breeds That Are “At Risk” for Producing a Foal With Neonatal Isoerythrolysisa
| Breed | At Risk for Aa | At Risk for Qa |
| Thoroughbred | 2% | 16% |
| Standardbred—pacer | 22% | b |
| Standardbred—trotter | 3% | b |
| American Saddlebred | 25% | 88% |
| Quarter Horse | 25% | 68% |
| Arabian | 3% | 72% |
aBased on the lack of all alleles that include factors Aa or Qa.
bMares all lack factor Qa and are technically “at risk,” but all Standardbred stallions also lack the factor.
such as nephrosis and centrilobular hepatic necrosis, may also be present.
■ Treatment and Prognosis In most cases, by the time the problem is recognized clinically (e.g., when the foal is about 24 hours of age), the bulk of colostral antibody will have been depleted from the mare’s milk and the absorptive ability of the foal’s gut will have diminished. Withholding milk at this point is of questionable benefit.
Stress should be minimized and exercise restricted. Affected foals have decreased exercise tolerance and can collapse and die if forced to follow their dams. Generalized supportive care should be administered as indicated by clinical parameters. Intravenous fluids are frequently indicated to promote diuresis to minimize the effects of the large hemoglobin load presented to the kidneys. Acid-base balance should be monitored and corrected if indicated.
If the anemia becomes severe (e.g., PCV = 10% to 15%), transfusions that provide RBCs should be considered. Unless the PCV drops below this level, transfusion may not be necessary if rest is enforced. The object of transfusion is to provide the affected foal with RBCs that will not be destroyed by the maternal alloantibodies that were absorbed from colostrum. The foal is immunologically naive and will not have autologous alloantibody directed against any RBCs that would have an immediate effect on transfused RBCs. Thus the key is to select an RBC donor whose cells will not be destroyed by the maternal antibody derived from colostrum. Selection of a donor with cells that will not be destroyed by the maternal antibody can be based on the results of crossmatching using mare serum, colostrum, and/or foal serum (all contain the same antibody) and the donor RBCs.
Washed RBCs from the dam are obviously the perfect choice in terms of cells that will not react with the alloantibodies present in the foal. Therefore, to avoid administering additional harmful antibody to the foal, the mare’s serum (containing antibody against the foal’s cells) must be removed by washing before administration. Up to 6 to 8 L of blood can be collected from the mare into containers with acid-citrate-dextrose (ACD) or sodium citrate (3.8% NaCitrate solution; 1 part NaCitrate/9 parts blood), although 3 to 4 L usually provide sufficient RBCs. The preparation of large volumes of washed cells is aided by a large-volume centrifuge, but the procedure can be accomplished by serial sedimentations. Anticoagulated blood from the mare is allowed to settle for 1 to 2 hours. The plasma is aseptically drawn off, a similar or greater volume of sterile isotonic saline (0.9% NaCI) is added to the RBCs and mixed, and the RBCs are again allowed to settle. The saline is then drawn off and discarded. At this point the RBCs can be resuspended in an equal volume of isotonic saline for administration, or the washing procedure can be repeated. The sedimentation method is less desirable than centrifugation because it is slower and does not remove as much of the offending antibody. The aim is to dilute any harmful antibody to insignificant levels.
If it is not possible to use the dam’s RBCs, alternative donors can be selected. The donor should lack the antigen to which the alloantibody is directed. Because it is generally not possible to blood type donors on short notice, a previously identified horse that has been determined by blood typing to be Aa-, Qa-, and free of alloantibody is a good choice for a donor
because most cases of NI are associated with these two antigens. The odds of randomly selecting a donor that lacks the offending type (e g., Aa-, Qa-, or both) and would therefore be a suitable donor vary significantly with the breed and would mirror the percentage of the population of mares at risk. For example, the odds of finding an Aa- Thoroughbred to serve as an RBC donor would be about 1 in 50 (2%), whereas in Quarter Horses the odds would be about 1 in 4 (25%) (see Table 53.12). The sire of the foal is not the donor of choice. He shares the same RBC antigens as the foal, and his cells will react with the maternal alloantibody present, adding more of a load to the foal's mononyclear phagocyte system as they are destroyed.
In the case of mules, the same considerations in the use of washed cells would be necessary if the dam's cells were used. However, because the offending antibody is generally directed against a unique donkey antigen, it appears that RBCs from any horse would be satisfactory. Horses do not appear to make naturally occurring antibodies against donkey factor; therefore in most cases it is not necessary to wash the cells from horses that would not be likely to have been immunized by pregnancy against donkey factor.
Transfusion of RBCs that will not react with maternal alloantibody results in the introduction of an obviously incompatible cell into the foal. Transfused cells may sensitize the foal to future transfusions, causing reactions (perhaps not within hours or a few days but potentially within a week). The RBCs will probably not survive for long periods in the circulation, and such transfusions should be considered temporary measures. These possibilities must be considered when weighing the potential good versus potential harm of such transfusions.
To produce clinical improvement, 1 to 4 L of washed RBCs or whole blood are usually adequate, although some cases may require repeated transfusion if the anemia progresses. Exchange transfusions can be done whereby blood is administered via one jugular and withdrawn simultaneously from the opposite jugular and discarded, allowing administration of large volumes of blood without overloading the vascular system. There is no good evidence to suggest that this is more effective than simply providing a source of RBCs that are unaffected by the maternal antibody.
Limited transfusion studies in adult horses have suggested that transfused RBCs do not survive long in circulation (e.g., 2 to 4 days), whereas in foals cells survived slightly longer (e.g., 4 to 6 days).48,49 PCVs in foals with NI commonly increase following transfusion and then gradually decline. This decline is probably not great cause for concern if it is gradual because the PCV of the foal generally levels off as the offending maternal antibody is metabolized. However, even short-term survival of cells may be of benefit in severely affected foals and may allow them to survive until the titer of maternal anti-RBC immunoglobulin has declined in the circulation. Hepatic failure in foals administered blood transfusions may be the result of iron intoxication. Administration of drugs that increase iron excretion, such as deferoxamine mesylate, are of potential value.50
The prognosis varies depending on the quantity of antibody ingested, the rapidity of onset of signs, and the degree of anemia. Peracute cases may die before the problem is recognized, with no chance for administration of therapy. Foals that develop the condition more slowly may respond to supportive care or transfusion if the PCV continues to fall.
■ Prevention and Control Several strategies are available for prevention of NI.51,52 First, identify broodmares at risk for development of NI by testing them for the presence of Aa and Qa. Mares negative for either antigen, which means they could potentially make antibodies against them, should be considered at risk. One subsequent strategy would be to breed at-risk mares to Aa-negative and Qa-negative stallions, thus eliminating the possibility of the foal inheriting the offending antigens. However, in breeds in which a relatively small part of the population is negative for these antigens, identifying a stallion that is negative for both antigens and suitable based on other criteria may be difficult. The percentage of at-risk females based on the presence or absence of Aa or Qa is somewhat balanced by the number of males able to transmit the offending antigen. For example, all Standardbred mares would be considered at risk because of the absence of Qa; however, because the antigen Qa is not present in the Standardbred population, no stallions have the Qa antigen to pass on to foals.
In the circumstance of unknown or incompatible matings, sera from at-risk mares should be screened for the presence of anti-RBC antibodies within 30 days before foaling. This can be done by submitting a serum sample and an anticoagulated sample to a screening laboratory. A panel of 10 to 12 different RBCs selected to represent all major blood groups is adequate to screen for anti-RBC antibody in the absence of blood from the sire. If results of serum testing are equivocal (e.g., low but positive titer, especially if there is anti-Aa or anti-Qa activity), the test should be repeated closer to the time of parturition because the levels of offending antibody can rise quickly late in gestation.
If anti-RBC antibody is detected in the mare before parturition, the colostrum should be checked for reactivity against the foal's RBCs before allowing the foal to ingest colostrum. An alternative source of colostrum should be provided to the foal. Most field screening tests of colostrum have not proved to be satisfactory for practical use; however, the jaundiced foal agglutination (JFA) test described in Box 53.2 has been shown to correlate well with the standard hemolytic assay, and it may detect antibody that does not react on the standard agglutination tests.7
Horses negative for Ca frequently make anti-Ca antibody; however, this antibody is not known to produce adverse effects in the foal. It has clouded the issue of field screening tests for NI because it commonly causes positive reactions in most screening tests at low dilutions. Because these tests do not differentiate between antibody to Aa, Qa, Ca, or any other blood group, anti-Ca antibody, when present in low dilutions, is responsible for many false-positive reactions. This antibody actually appears to play a protective role in the prevention of sensitization of mares to NI through a mechanism of antibody-mediated immune suppression. Aa- and Qa- mares that are also Ca- (and thus often produce anti-Ca antibody) become sensitized to Aa and Qa at a significantly lower rate than mares that are Ca+.53 This is attributed to the fact that anti-Ca antibody is produced by Ca- mares. The anti-Ca antibody may more rapidly remove potentially sensitizing cells from the circulation before they stimulate production of antibody against Aa or Qa.
Uncommonly, other antigens have been associated with NI in foals. These include factors Db, Dc, Dg, Ua, Pa, Qc, and Ab.2,4,53-58 Two other factors, R and S, have been described to be associated with NI, but they are not detected by routine hemolytic or agglutinating methods and are detected only using an antiglobulin test (direct Coombs test).41 Because of the difficulty in testing for these antigens, there has never been sufficient agreement among laboratories to allow international designation. These antigens may not be detected using the JFA test.25 Estimates are that 1 in 2000 pregnancies may result in sensitization against some other antigen besides Aa or Qa. Because these cases occur so infrequently, it is not practical to consider mares without these antigens to be at risk for NI. A blood-type evaluation of mares with a history of production of foals with NI should be done to identify the offending antibody or antigen.
A list of laboratories providing NI screening tests (e.g., Aa, Qa, and Ca typing and screening of sera for alloantibody) is provided in Box 53.3. Routine agglutinating crossmatch tests using mare serum or colostrum and foal or sire cells
■ BOX 53.2
■ BOX 53.3
Jaundiced Foal Agglutination (JFA) Test
Materials
1. Centnfuge capable of spinning 300? to 600? gravity
2. Test tube rack
3. Test tubes: 13- ? 100-mm disposable or blood collection tubes
4. Pasteur pipettes and rubber bulbs or other pipette system to deliver 1-mL volumes
5. Isotonic (0.9%) saline at room temperature
6. Serum or colostrum from the mare; RBCs from mare and foal, preferably in EDTA anticoagulant
Methods
1. Collect colostrum from the mare.
2. Collect an EDTA-anticoagulated blood sample from the foal before nursing.
3. Set up six tubes and add 1 mL of saline to each tube. Label the tubes Control,1 : 2, 1 : 4, 1 : 8, 1 : 16, and 1 : 32.
4. Make serial dilutions of the colostrum at 1: 2, 1: 4, 1: 8, 1 : 16, and 1: 32 in five of the tubes. Add 1 mL of colostrum (or serum) to the tube labeled 1 : 2 and mix. Then take 1 mL from that tube and add it to the second tube labeled 1:4 and mix. Take 1 mL from that tube and add it to the third tube labeled 1 : 8, and so on, until all five dilution tubes have been filled. Discard 1 mL from the last tube labeled 1: 32.
5. Add 1 drop of the foal's whole blood to each of the six tubes and mix.
6. Centrifuge the tubes for 2 to 3 minutes at a medium speed (300? to 500? gravity).
7. Invert each tube, pouring out the liquid contents; observe the status of the button of RBCs at the bottom of the tube. Complete agglutination causes the cells to remain tightly packed in the button; strong agglutination causes the cells to remain in large clumps; for weaker agglutination the cells are in smaller clumps as they run down the side of the tube. When there is no agglutination, the cells easily flow down the side of the tube. This should be the case in the control tube. If the cells in the control tube are clumped, they may be autoagglutinating and the results will have questionable validity.
8. If there is a positive reaction with the foal's cells, the test should be run with the dam's own RBCs to be certain that neither the conditions of the test nor the viscosity of the colostrum is causing the agglutination.
9. Positive reactions at 1:16 or greater are considered significant. At levels of 1:16 or greater, this test correlates well with the standard hemolytic assay. At dilutions of less than 1:16, the correlation is not as good, and more falsepositive results will be recorded. Also, other factors such as the viscosity of colostrum make less diluted samples more difficult to read.
EDTA, Ethylenediaminetetraacetic acid; RBCs, red blood cells.
can be performed by most veterinary or human hematology laboratories.
Ruminants
NI is not a naturally occurring disease in cattle, sheep, or goats. Its occurrence in cattle has been associated with administration of vaccines derived from blood origin, such as certain anaplasmosis and babesiosis vaccines.2 When these vaccines are used on breeding females, they may sensitize the dam to certain blood groups, most commonly in the A and F systems. Under chance circumstances, if the blood types of the sire and offspring reflect these systems and the dam has produced alloantibodies, an isoimmune hemolytic crisis may appear in the calf associated with successful passive transfer.
Laboratories Providing Equine Neonatal Isoerythrolysis Testinga in North America
Clinical Laboratory Services-Hematology University of California, Davis
Veterinary Medical Teaching Hospital Central Laboratory Receiving, Room 1033 One Garrod Drive
Davis, CA 95616-8747 (530) 752-8684 www.vetmed.ucdavis.edu/vmth Animal Genetics Testing and Research Laboratory 108 Gluck Equine Research Center
University of Kentucky Lexington, KY 40546-0099 (859) 218-1212 www.ca.uky.edu/gluck/AGTRL.asp Rood and Riddle Veterinary Laboratory Rood and Riddle Equine Hospital 2150 Georgetown Road
Lexington, KY 40511 (859) 233-0331 www.roodandriddle.com/laboratory.html Hagyard Laboratory
Hagyard Equine Medical Institute 4250 Iron Works Pike
Lexington, KY 40511-8412 (859) 259-3685 www.hagyard.com/Hagyard-Laboratory.html
aSerum or colostrum from the suspect mare is required to screen for the presence of alloantibody. Acid-citrate-dextrose anticoagulated blood is generally preferred for screening for red blood cell antigens.
Hemolytic crises are rare in sheep and are even difficult to induce experimentally.2
Bovine neonatal pancytopenia (BNP) is a fatal condition reported in Europe and New Zealand in a subset of neonatal calves born to cows vaccinated with a certain vaccine product (Pregsure® BVD [Pfizer, New York, N.Y.]); the vaccine has been withdrawn from the market.59 Nevertheless, cases of BNP continue to be seen in calves born to dams that have been historically vaccinated. This vaccination-induced alloimmune condition is characterized by marked anemia, thrombocytopenia, lymphopenia, and bone marrow hypoplasia, causing severe internal and external hemorrhage. The involvement of colostrum was identified by inducing the disease in newborn calves (about 10 days when offering colostrum from vaccinated cows) and by preventing disease by colostrum substitution.60 Studies demonstrated that the alloantibodies produced by vaccinated cows reacted with proteins from the cell line used to manufacture the vaccine. One study demonstrated that the development of BNP involves a heritable trait (19%) determined by the dams; that is, genetic differences between dams explain in part why the colostrum of some but not all vaccinated dams cause BNP in the calf or cause subclinical disease.61 Genetic inheritable differences do not seem to involve major histocompatibility complex classes I and II genes or differences in Ig isotypes but rather unknown genes that control the amount of alloantibody produced by the vaccinated dams.62,63
Neonatal Alloimmune Thrombocytopenia
■ Equine
Definition and Etiology
Neonatal alloimmune thrombocytopenia (NAIT) is a condition characterized by the destruction of platelets in the circulation of a foal by alloantibodies of maternal origin absorbed from colostrum.63 The syndrome has been observed in horse and mule foals.44,64,65 The prevalence of this condition is unknown. Some cases may be asymptomatic, and the condition may be self-limiting as alloantibody is metabolized.
■ Clinical Signs and Differential Diagnoses Few clinical signs may appear unless foals are traumatized. Affected foals may have prolonged bleeding from venipuncture sites. Petechial hemorrhages may or may not be present.64 Other conditions often associated with thrombocytopenia in neonates include sepsis, disseminated intravascular coagulation, equine infectious anemia, drug-induced thrombocytopenias, and angiopathies. The challenge is to determine whether the thrombocytopenia is primarily due to allogeneic antibodies or secondary to some other disease process.
■ Clinical Pathology Profound thrombocytopenia in the absence of other hematologic changes typifies uncomplicated NAIT. Evidence of successful passive transfer is present based on quantification of serum IgG. Affected foals are thrombocytopenic. Thrombocyte counts less than 10,000 cells∕μL have been observed.64
Demonstration of significant amounts of antibody in the colostrum (or serum from the mare) that are directed against platelet antigens expressed by the foal provides a definitive diagnosis of NAIT; however, assays for equine platelet-bindable and platelet-associated immunoglobulins are not routinely available.
■ Pathophysiology The pathophysiology is believed to mirror that of NI. In mares sensitized to platelet antigens, alloantibodies are concentrated in the colostrum late in gestation. These antibodies are passively transferred to the foal. If the foal's platelets carry the antigen that the antibody recognizes, the platelets become antibody coated. Subsequently they are removed prematurely by the mononuclear phagocyte system. Platelet antigens have not been characterized in horses, although platelet-associated antibodies have been recognized in affected foals. Circulating antibody is removed by attachment to platelets and rapid clearance by the mononuclear phagocyte system.
■ Treatment and Prognosis Circulating antibody is removed relatively quickly, and treatment may not be necessary. Platelet-rich plasma may be indicated in cases of severe thrombocytopenia accompanied by clinical signs of bleeding complications; however, as offending antibody is removed, the problem will tend to be self-limiting.
■ Prevention and Control Currently there are no screening tests to predict NAIT. Following production of an affected foal by a mare, it would be prudent to provide an alternate source of colostrum to foals born in subsequent pregnancies.
A rare condition of severe thrombocytopenia, neutropenia, and ulcerative dermatitis has been described in equine neonates, and clinical signs suggest an association with passively transferred colostral antibodies or other factors.66
Alloimmune Neonatal Neutropenia
■ Equine
Definition and Etiology
Alloimmune neonatal neutropenia (ANN) is a condition characterized by the destruction of neutrophils in the circulation of a foal by alloantibodies of maternal origin absorbed from colostrum.67,68
■ Clinical Signs and Differential Diagnoses In the absence of secondary bacterial infections, no clinical signs are observed. Diagnosis depends on the presence of persistent neutropenia in the absence of evidence of increased consumption or decreased production of neutrophils.
■ Clinical Pathology Persistent neutropenia in the face of normal clinical conditions has characterized reported cases.
■ Pathophysiology The pathophysiology is believed to mirror that of NI. In mares sensitized to granulocyte antigens, alloantibodies are concentrated in the colostrum late in gestation. These antibodies are passed to the foal through passive transfer. If the foal's neutrophils carry the antigen that the antibody recognizes, the neutrophils become antibody coated. Subsequently they are removed prematurely by the mononuclear phagocyte system. Neutrophil antigens have not been characterized in horses, but neutrophil-associated antibodies have been recognized in affected foals. Circulating antibody is removed by attachment to neutrophils and rapid clearance by the mononuclear phagocyte system.
■ Treatment and Prognosis ANN is self-limiting in infants and typically resolves over several weeks as maternal antibodies diminish. Administration of antimicrobial drugs may provide protection against bacterial infections. Corticosteroids are not currently recommended for treatment of the condition in human infants and have not been used in equine cases. Administration of recombinant human granulocyte colonystimulating factor (rhG-CSF) was associated with increases in neutrophil counts in one affected foal.67
■ Prevention and Control Demonstration of agglutinating neutrophil antibodies or immunoglobulin coating of neutrophils using flow cytometry from the foal may be used to confirm a diagnosis.67 Following production of an affected foal by a mare, it would be prudent to provide an alternate source of colostrum to foals born in subsequent pregnancies.