Epidemiology of Histoplasmosis
The main historical facts and pioneering events that contributed to our knowledge on epidemiology of human and animal histoplasmosis are summarised in Fig. 5.2. A large diversity of domestic and wild mammals can be infected by H.
capsulatum (Chermette and Guillot 2010): non-human primates, equids (including horses, donkeys and mules), cattle, dromedaries, rabbits (Brandoa et al. 2014), hedgehogs (Snider et al. 2008), pigs, dogs, cats, other carnivores such as grey (Urocyon cinereoargenteus) and red foxes (Vulpes vulpes), brown bears (Ursus arctos), raccoons (Procyon lotor), striped (Mephitis mephitis) and spotted (Spilogale putorius) skunks, European badgers (Meles meles) and sea otters (Enhydra lutris) (Morita et al. 2001). Spontaneous or experimentally induced histoplasmosis has also been reported in various species of rodents among which common (Mus musculus) and white-footed mice (Peromyscus leucopus); black (Rattus rattus), grey (Rattus norvegicus) and spiny rats (Proechimys semispinosus); and opossums of the genera Didelphis and Philander. Histoplasma organisms have also been isolated from many different species of bats whose guano is considered the best substrate for the proliferation and survival of the filamentous form of the subspecies capsulatum.Inhalation is supposed to be the primary mode of entry of H. capsulatum var. capsulatum. Animals that are immunocompromised or were subjected to a large dose of infectious fungal elements are at a greater risk to develop infection with dissemination. Infection is caused by the inhalation of propagules from the saprobic
OO
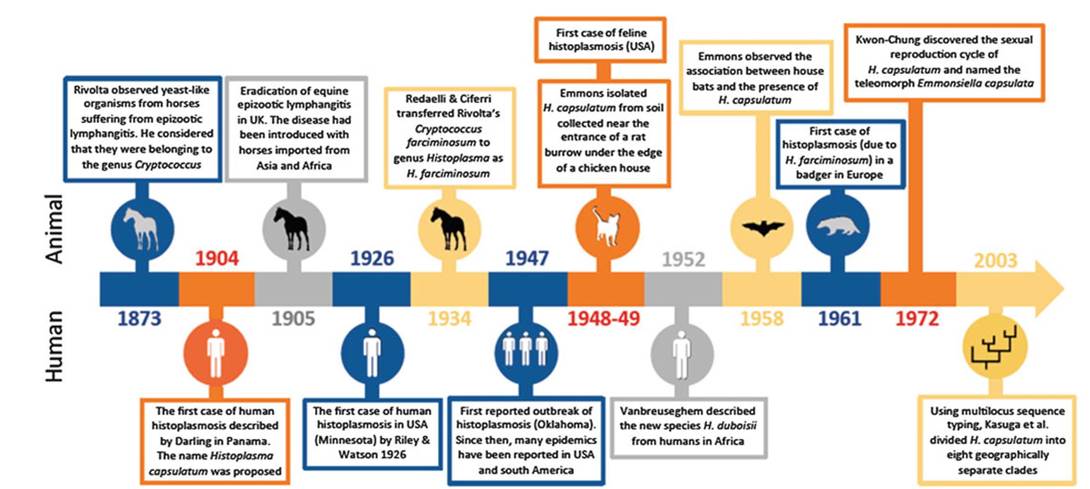
Fig. 5.2 Chronological order of the main facts and pioneering events that contributed to the epidemiological studies on human and animal histoplasmosis
filamentous phase.
This phase develops in special habitats, particularly on bat guano that has accumulated in confined spaces such as caves. A second possible mode of entry by ingestion of infective material has been suggested. This could account for cases of primary gastrointestinal histoplasmosis in dogs and cats (Bromel and Sykes 2005; Stark 1982). A moderate climate with constant humidity seems to be the most appropriate combination for the development and survival of Histoplasma. In companion animals, infection due to H. capsulatum var. capsulatum is most frequently observed in endemic/enzootic regions of the Mississippi and Ohio River valleys in North America (Bromel and Sykes 2005; Sykes and Guillot 2015). Cases were also reported from South America (Forjaz and Fischman 1985), Italy (Mantovani et al. 1968; Reginato et al. 2014), Greece (Mavropoulou et al. 2010), Japan (Murata et al. 2007; Ueda et al. 2003) and Australia (Mackie et al. 1997). Recently, cases of feline histoplasmosis were reported in Colorado, California, New Mexico and Texas, locations that were traditionally considered as non-enzootic (Balajee et al. 2013). A massive H. capsulatum infection in juvenile raccoons from northern California was reported by Clothier et al. (2014). In 2005, an infected northern sea otter (Enhydra lutris kenyoni) was found on Kodiak Island, Alaska (Burek-Huntington et al. 2014). Histological examination revealed the presence of Histoplasma yeasts and the subspecies capsulatum could be identified by direct sequencing. The authors suggested that the sea otter was contaminated by migratory birds or through aerosol transmission.Europe is usually considered non-endemic/enzootic for H. capsulatum. However, in a review about humans with histoplasmosis in Europe over a 5-year period (from January 1995 to December 1999), Ashbee et al. (2008) identified eight patients (from Germany, Italy and Turkey) who had never travelled abroad and hence may correspond to autochthonous cases. Histoplasmosis has also been reported in wild and domestic animals in Europe.
Several investigations demonstrated that H. capsulatum may be responsible for cutaneous lesions in Eurasian badgers (Meles meles) in Switzerland (Burgisser et al. 1961), Denmark (Jensen et al. 1992), Germany (Grosse et al. 1997; Eisenberg et al. 2013) and Austria (Bauder et al. 2001). Infection was limited to the skin and subcutaneous lymph nodes. The badger's habitat and its rummaging and omnivorous mode of life are potential predisposing factors to infectious diseases, including tuberculosis and histoplasmosis. Recently, H. capsulatum var. capsulatum DNA was detected from the lungs of a bat (Nyctalus noctula) trapped in France (Gonzalez-Gonzalez et al. 2013). Disseminated histoplasmosis was described in a cat in Greece (Mavropoulou et al. 2010) and in dogs in Italy (Mantovani et al. 1968; Reginato et al. 2014).The subspecies farciminosum was epidemiologically investigated between January 2003 and June 2004 in 19,082 carthorses in Ethiopia (Ameni 2006). A mean prevalence of 18.8% was reported. The highest prevalence (39%) was observed in the Mojo region. The prevalence of infection was associated with average annual temperatures rather than mean annual rainfall. Statistically significant association was also observed between the altitude and the prevalence of infection: histoplasmosis was more frequently reported in humid and hot regions with an altitude from 1500 to 2300 m (Ameni 2006). Equine histoplasmosis has been reported from China, India, Indonesia, Iraq, Israel, Japan, Pakistan and Syria. In Africa, cases are reported in many countries: Algeria, Angola, Cameroon, Chad, Egypt, Ethiopia, Ghana, Morocco, Nigeria, Togo, Tunisia, Senegal and Sudan (Guerin 2010). Official disease distribution maps from OIE (Office International des Epizooties) indicate that equine histoplasmosis in Africa is restricted to Ethiopia, Senegal and South Africa (OIE WAHID Maps 2005). The disease is also reported from South and Central America (Al-Ani 1999). Compared to major equine epizootic diseases, infection due the subspecies farciminosum is not frequently diagnosed.
The confusion with other infectious diseases may probably account for this situation.Infections limited to the skin were also reported in cats residing in Switzerland (Fischer et al. 2013) and in eastern France (Fantini et al. 2014). In these cases, histopathological examination revealed the presence of microorganisms consistent with Histoplasma (Fig. 5.1), and the subspecies farciminosum was identified by PCR and MLST approach directly from the tissues (Guillot et al. 2015).
Inoculation is considered the primary mode of entry of H. capsulatum var. farciminosum, probably mostly by contamination of cutaneous wounds in horses (Guerin 2010). Nevertheless experimental transmission of the disease by subcutaneous or intradermal inoculation of pus containing H. capsulatum var. farciminosum yeasts gave inconstant results. Some cases of direct transmission between infected and healthy wounded animals or after mating have been suspected (Al-Ani and Al Delaimi 1986). The subspecies farciminosum has also been isolated from the digestive tract of hematophagous flies (Gabal and Hennager 1983). Ameni and Terefe (2004) indicated that there was a significant association between histoplasmosis and the presence of ticks in mules. Seasonal changes may have a significant effect upon direct transmission efficiency, as rain increases mud projections on wounds, delays healing processes and increases the risk of infection in horses (Guerin et al. 1992). Telluric contamination of exposed and vulnerable body areas such as limbs, eyes or nostrils could explain the main localisations of primary lesions.
5.3