Examination of Cranial Nerves
Examination of the cranial nerves is most easily carried out by examining the functions of groups of nerves that innervate particular regions of the head rather than by performing the examination in a strictly numeric order.
For example, examination of vision and other ocular functions such as the menace response, pupillary light reflexes, and physiologic nystagmus reveals the integrity of several cranial nerves, not only II, III,
FIG. 8.7 Muscle atrophy in the gluteal muscles in a horse with equine protozoal myeloencephalitis.
IV, and VI but also V (sensory to the cornea), VII (motor to the eyelids), and VIII (providing vestibular input to control the functions of III, IV, and VI), as well as centers within the brain.
CRANIAL NERVE I—OLFACTORY NERVE. Reliable and specific testing of the sense of smell is difficult in animals. Large animals require an intact sense of smell to eat properly, so it can be inferred that animals with good appetites possess an adequate sense of smell. Having a blindfolded animal track food moved from side to side in front of the nose may be helpful. Irritating substances such as ammonia should not be used for evaluation of olfactory nerve function; such compounds stimulate nociceptors in the nasal mucosa, which are the dendrites of the maxillary nerve (cranial nerve V), rather than stimulating olfactory receptors innervated by the olfactory nerve. Loss of the sense of smell is more likely to be caused by disease within the nasal passages than by a primary neurologic disease.
CRANIAL NERVE II—OPTIC NERVE. Vision and light perception are the functions of cranial nerve II, the optic nerve. Observing the animal's response to its environment provides a good initial assessment. Does it respond to visual cues such as movement or does it walk into objects? Noise may cause the animal to turn its head toward the sound, so the observer must be careful to distinguish such responses from those made in response to visual cues.
A maze can be set up using straw bales or other objects, and then the animal can be gently driven■ TABLE 8.6
Guide to Neuro-Ophthalmologic Lesion Location
| Lesion Location | Menace Response | Pupillary Light Reflex | Maze Test | ||
| Ipsilateral | Contralateral | Ipsilateral | Contralateral | ||
| Unilateral retina, optic nerve | Absent | Present | Absent (direct), slightly dilated | Normal | Abnormal |
| Bilateral retina, optic nerve, | Absent | Absent | Fixed | Fixed | Abnormal |
| optic chiasm | |||||
| Unilateral oculomotor nerve | Absent | Present | Dilated nonresponsive | Normal | Normal |
| Unilateral occipital cortex | Present | Absent | Normalb | Normalb | Abnormal |
| Bilateral occipital cortex | Absent | Absent | Normalb | Normalb | Abnormal |
| Bilateral vagosympathetic trunk | Present | Present | Miotic | Normal | Normal |
| Bilateral cerebellar cortex | Absent | Absent | Normal | Normal | Normalc |
aIn unilateral cases, assumes that contralateral eye is blindfolded.
bAssuming that no cortical swelling has occurred.cAnimals walk abnormally but recognize and generally avoid obstacles.
through the maze. Normal animals will avoid colliding with objects. Animals that are severely obtunded, however, may walk into objects even when they can see. Thorough evaluation of the complete neurologic examination is necessary to distinguish this from true blindness.
The menace response measures the integrity of the entire visual pathway. The afferent pathway runs from the retina via the optic nerves and internal capsule to the visual areas in the occipital lobe of the cerebrum. Information from the visual cortex is processed and relayed to the ipsilateral motor cortex. The efferent pathway of the menace response runs from the motor cortex via the pons to the nucleus of the facial nerve in the medulla oblongata and thence via the facial nerve to the orbicularis oculi muscle. It is not known whether this pathway passes through the cerebellum, but it is clear that the ipsilateral cerebellar cortex is required for a competent menace response. The menace test is performed by rapidly advancing the hand toward the eye and observing closure of the eyelid. In addition to closure of the eyelid, some animals display a generalized avoidance response characterized by coordinated movement of the head and neck away from the stimulus. The opposite eye may be covered to ensure that only one eye is being stimulated. Care must be taken not to touch the face or eyelashes or create air currents near the face. Separate menacing gestures must be directed from both the nasal and the temporal aspects, in order to cover the entire visual arc. The menace response measures the integrity of the retina, optic nerve, optic chiasm, optic tracts, thalamus, internal capsule, and occipital cortex. There is approximately 90% crossing over of optic nerve fibers in the optic chiasm of livestock. Unilateral lesions central to the optic chiasm—in the optic tract, internal capsule, thalamus, or occipital lobe—cause loss of vision in the opposite eye, with apparently normal vision in the ipsilateral eye.
It should be noted that normal neonatal animals may not display a response to a menace gesture because this is a learned response.Menace deficit may be the result of facial nerve paralysis. In such cases the animal does not blink but shows avoidance of the stimulus by pulling the head away. Facial nerve deficits will be apparent in these animals by their inability to close the eyelids in response to cutaneous stimulation around the lid margins (palpebral reflex) and by other signs such as drooping of the lip and ear on the same side. Animals with cerebellar disease may also display a menace deficit yet possess normal vision. Menace deficits resulting from facial nerve or cerebellar disease may be differentiated from deficits in other areas by maze testing. Animals with cerebellar or facial nerve disease retain visual acuity and maneuver through the course successfully. In addition to the optic pathways, the test measures the integrative pathways in the frontal and parietal lobes of the brain, the motor neurons, and the proprioceptive pathways (Table 8.6). Myasthenic diseases (e.g., botulism, hypocalcemia, hypomagnesemia) result in bilaterally decreased menace and palpebral responses but do not produce blindness.
The pupillary light reflex measures the integrity of the retina, optic nerves, optic chiasm, pretectal and oculomotor nuclei in the midbrain, oculomotor nerve, ciliary ganglia, and constrictor pupillae muscle. The test is performed by shining a bright light into each eye and observing constriction of the pupil in the ipsilateral eye (direct reflex) and the contralateral eye (indirect reflex). Lowering the ambient light level may facilitate this test by causing the pupils to dilate. The reflex in large animals is considerably slower than those observed in cats and dogs. A “swinging light” test has been recommended in large animals to reduce the dazzle and startle responses elicited by suddenly shining a bright light into the eyes.46 From a position directly in front of the animal, a strong light is slowly swung from one eye to the other while bringing the light closer and closer to the head and observing the responses in each pupil.
The effects on the pupillary light reflex of lesions at various levels along the visual pathway are shown in Table 8.6.Unilateral lesions in the forebrain may result in blindness of the opposite eye. With forebrain lesions, the pupillary light reflexes are usually normal. If the forebrain disease is accompanied by increased intracranial pressure, the oculomotor nerve or nucleus may be compressed in the midbrain, resulting in ipsilateral mydriasis.
CRANIAL NERVES III, IV, AND VI-OCULOMOTOR, TROCHLEAR, AND ABDUCENT NERVES. The position of the globe in the orbit is governed by the activities of the oculomotor, trochlear, and abducent nerves. Dysfunctions of these nerves result in deviation of the globe that is constant in all head positions. Loss of oculomotor nerve function results in ventrolateral strabismus. Trochlear nerve dysfunction results in rotation of the dorsal aspect of the globe away from the midline (dorso- medial strabismus). Loss of abducent nerve function results in medial strabismus and inability to retract the globe, which is best demonstrated by closing the eyelid and then pushing on the globe through the eyelid. The normal reflex is a retraction of the globe.
Competent function of the oculomotor, trochlear, and abducent nerves is also required for the oculocephalic reflexes. When the head is turned from side to side in a horizontal plane, physiologic horizontal nystagmus is observed, with the fast phase of the nystagmus in the direction of head movement. The sensory receptors for this reflex lie within the semicircular canals of the inner ear, and they detect angular acceleration of the head. Input from the semicircular canals is transferred to the vestibular centers in the medulla oblongata and the cerebellum and then to the nuclei of cranial nerves III, IV, and VI. Lesions of the peripheral or central components of the vestibular system can also result in abnormal eye position (strabismus) and movement (nystagmus), described in more detail later.
With peripheral or central vestibular lesions, however, eye position typically changes when the head and neck are moved, in contrast to the constant deviation of the globe seen with direct lesions to the oculomotor, trochlear, and abducent nerves.The oculomotor nerve is motor to the levator palpebrae superioris muscle, the most important of the muscles responsible for elevation of the upper eyelid. Lesions of the nerve cause ptosis (drooping of the upper eyelid) in addition to the signs described previously. Ptosis can also be induced by lesions of the sympathetic nerve supply to the eye because of paralysis of the superior tarsal (Muller’s) muscle and, in the horse only, by lesions of the facial nerve, which innervates the levator anguli oculi medialis muscle (see later).50
CRANIAL NERVE V—TRIGEMINAL NERVE. The trigeminal nerve is sensory to the face and motor to the muscles of mastication. The sensory functions of the trigeminal nerve are tested by lightly stimulating the forehead, upper and lower jaws, and muzzle using the tip of a closed hemostat or a finger. The normal response is one of avoidance using neck, facial, and appendicular musculature; the animal usually pulls the head away and blinks simultaneously. Some areas such as the cheeks, forehead, and chin are normally less sensitive, whereas the periorbital region, nasal planum, nasal septum, and lips are sensitive. The test evaluates the function of the sensory part of the trigeminal nerve, the trigeminal ganglion, the nucleus and spinal tract of the trigeminal nerve, the pontine sensory tract nucleus of cranial nerve V, the thalamus, and the sensorimotor cortex. After the trigeminal nerve enters the lateral aspect of the pons, axons both ascend and descend through the medulla as the spinal tract of the trigeminal nerve. Ascending information ultimately reaches the sensorimotor cortex via the thalamus, where it is consciously perceived. Descending information projects to the nucleus of the facial nerve in the medulla and also into the first cervical spinal segment. Unilateral loss of facial perception of touch most commonly results from damage to the peripheral portion of the trigeminal nerve, the trigeminal ganglion in the petrosal bone of the skull, or the contralateral forebrain (i.e., thalamus or cerebrum). Lesions affecting the spinal tract of the trigeminal nerve in the medulla and midbrain would likely be fatal because they also would affect adjacent respiratory and cardiovascular centers in the brainstem. Patients with bilateral facial hypalgesia most often have bilateral forebrain disease.
The palpebral reflex is elicited by lightly touching the periorbital area (both above and below the eye) and observing a brisk closure of the eyelid. This reflex reflects the sensory function of the trigeminal nerve and the motor function of the facial nerve and orbicularis oculi muscle. Simultaneous loss of the menace response and the palpebral reflex suggests a lesion in the facial nerve or the orbicularis oculi muscle. Loss of the palpebral reflex with normal menace responses suggests a lesion in the trigeminal nerve or ganglion. Loss of menace response with preservation of the palpebral reflex and pupillary light reflex suggests the presence of either contralateral forebrain disease or ipsilateral cerebellar cortical disease.
The jaw should be opened to assess the strength of the masticatory muscles. This measures both the sensory (proprioceptive) fibers of the trigeminal nerve and the motor component of the nerve. Bilateral lesions of the motor component of the trigeminal nerve result in a dropped jaw. Affected animals may protrude the tongue but can retract it normally when stimulated. Animals with dropped jaws may drool because they cannot hold saliva within the oral cavity. Unilateral lesions of the trigeminal nerve produce asymmetric jaw closure, with deviation of the mandible away from the affected side, and neurogenic atrophy of the visible masticatory muscles (i.e., masseter and temporalis).
CRANIAL NERVE VII—FACIAL NERVE. The motor nucleus of cranial nerve VII (the facial nerve) originates in the middle and ventral part of the medulla oblongata. The motor fibers are distributed to muscles of facial expression. Just as the motor fibers are exiting from the lateral aspect of the brainstem, they merge with axons from the parasympathetic facial nucleus. These autonomic fibers innervate the lacrimal and salivary glands and separate from the motor component of the facial nerve as it traverses the petrous temporal bone. Lesions of cranial nerve VII located between the brainstem and this separation usually result in “dry eye.” More distal lesions, however, have no effect on tear production. The tone of the facial musculature is examined by palpation of the ears, lips, eyelids, and muzzle. Clinical signs of facial nerve dysfunction include drooped ear and lips, drooling saliva, and retention of food in the cheek pouch on the denervated side (Fig. 8.8). Closure of the eyelids is weak in partial facial nerve lesions and absent in severe lesions. Despite this, there is slight drooping of the upper eyelid (ptosis) because of paralysis of the levator
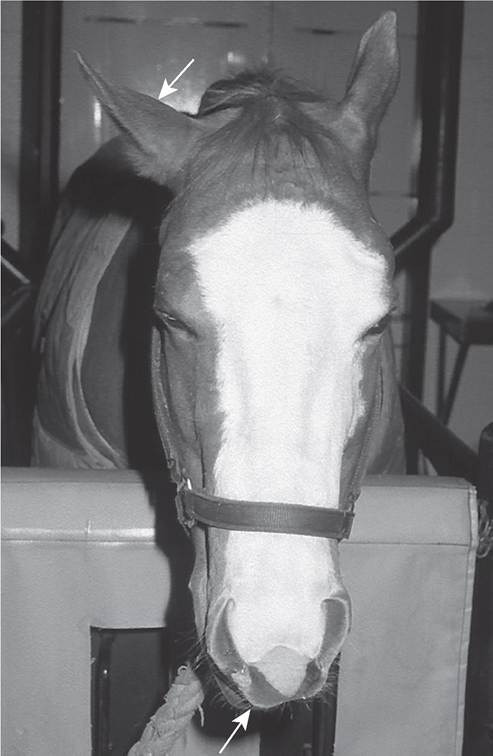
FIG. 8.8 Acute right facial paralysis in a horse with guttural pouch mycosis. Note the drooped right ear and deviation of the muzzle toward the left side (arrows).
anguli oculi medialis muscle, which contributes to eyelid retraction. In species with a soft muzzle (e.g., horses, sheep, goats), there is a marked deviation of the nasal filtrum away from the side with the lesion after unilateral loss of facial nerve function. The filtrum of affected cattle is not deviated because of the large amount of fibrous tissue in the planum nasale. In chronic facial paralysis the face may be deviated toward the affected side because of atrophy and contracture of the denervated musculature of the face. In horses neurogenic atrophy of the parotidoauricularis muscle is evident as a groove behind the vertical ramus of the mandible.
CRANIAL NERVE VIII-VESTIBULOCOCHLEAR NERVE
Vestibular System. The function of the vestibular system—which is composed of the sensory structures in the inner ear (semicircular canals, utriculus, saccule), the vestibular portion of cranial nerve VIII, and the central components of the vestibular system in the medulla oblongata and cerebellum—is tested by assessment of gait, extensor tone, head posture, and eye movements. Signs of vestibular dysfunction include a staggering gait, circling, falling, rolling, head tilt, and spontaneous nystagmus. Lesions can be classified as peripheral or central. Diseases affecting the inner ear or cranial nerve VIII result in signs of peripheral vestibular disease. Lesions affecting vestibular structures in the medulla oblongata result in central vestibular signs, and lesions affecting vestibular structures in the cerebellum result in paradoxical vestibular signs. Blindfolding affected patients results in a worsening of clinical signs because of elimination of visual compensatory mechanisms. Recumbent animals with vestibular lesions tend to lie with the side of the vestibular lesion downward. When turned, these animals spontaneously rotate back to the lesion-down position and may strongly resist attempts to turn them over. Animals with unilateral vestibular disorders typically have ventral strabismus in the ipsilateral eye and dorsal strabismus in the contralateral eye. Bilateral, symmetric lesions of the vestibular system are rare. Bilateral vestibular dysfunction does not cause head tilt, nystagmus, or strabismus. Affected animals are reluctant to move. They stand with a basewide posture, with the head held low, and fall easily when forced to move.
CRANIAL NERVES IX, X, AND XI-GLOSSOPHARYNGEAL, VAGUS, AND SPINAL ACCESSORY NERVES. Cranial nerves IX, X, and XI originate in the nucleus ambiguus, a column of motor neurons in the medulla oblongata. They provide motor control to the muscles of the neck, pharynx, and palate. The vagus nerve contains efferent fibers that stimulate the secretions of glands of the visceral and respiratory mucosa and control forestomach motility in ruminants. The glossopharyngeal and accessory nerves carry afferent fibers from the mucosa of the tongue, larynx, and pharynx.
The signs of glossopharyngeal and vagus nerve dysfunction include dysphonia (roaring, snoring), dysphagia, and regurgitation. Animals with pharyngeal paralysis regurgitate food and saliva from the nose. Roaring is a characteristic stridor emanating from the larynx. The abnormal sound may be heard only during exercise. Functional examination of these nerves should include auscultation of the larynx for stridor, observation of the animal as it swallows, passage of a nasogastric tube to evaluate deglutition, endoscopic examination to evaluate pharyngeal and laryngeal movements, and palpation of the larynx for muscle atrophy. The slap test, described earlier, is a test for function of the vagal innervation of the larynx via the recurrent laryngeal nerve. Specific descriptions of the endoscopic appearance of pharyngeal paralysis and roaring are presented elsewhere (see Chapter 31).
Signs of spinal accessory nerve dysfunction are extremely rare and include atrophy of the trapezius, sternocephalicus, and brachiocephalicus muscles.
SYMPATHETIC INNERVATION OF THE HEAD—HORNER'S SYNDROME. Preganglionic sympathetic motor fibers that innervate structures of the head originate from the first three thoracic spinal cord segments. These fibers ascend the neck in the vagosympathetic trunk to the cranial cervical ganglion under the tympanic bulla, where they synapse with postganglionic sympathetic fibers. The postganglionic fibers are distributed to the smooth muscles of the head via the ciliary nerves, passing through the petrous temporal bone area. Lesions anywhere along the course of the preganglionic or postganglionic sympathetic nerves, in spinal cord segments T1 to T3, or, rarely, in the upper motor neuron component of the sympathetic pathway in the cervical spinal cord or brainstem (tectotegmentospinal tract) cause a characteristic constellation of clinical signs known as Horner’s syndrome. Signs include miosis, ptosis, and increased warmth on the ipsilateral side of the face. In cattle there is a loss of sweating on the ipsilateral half of the planum nasale, whereas in horses there is excessive sweating on the affected side of the head and upper neck. Miosis is produced by lack of sympathetic tone to the affected pupil. Diseases that could produce Horner's syndrome in large animals include lesions of the T1 to T3 spinal cord segments, mediastinal or thoracic abscesses or neoplasia, trauma or infection involving the cervical sympathetic trunk, esophageal perforations, guttural pouch mycosis in horses, otitis media and interna, and retrobulbar abscesses. Lesions of the midbrain may produce miotic pupils without other signs of Horner's syndrome.
CRANIAL NERVE XII—HYPOGLOSSAL NERVE. The hypoglossal nerve supplies motor innervation to the muscles of the tongue and the geniohyoideus muscle. The nucleus is located in the caudal medulla oblongata. Hypoglossal nerve function is tested by pulling the tongue out of the mouth. Normal animals should have forceful resistance to passive manipulation of the tongue. Lesions of the hypoglossal nerve result in flac- cidity and atrophy of the tongue. Acute lesions result in deviation of the tongue away from the side of the lesion; with time the tongue may deviate toward the affected side because of muscle atrophy and contracture.