Expansion Due to Fungal Novelties
In addition to changes in opportunity, also fungal populations change perpetually in their genetic makeup. For instance, virulent genotypes may be novel, or at least the hosts are confronted with fungal genotypes to which they lacked resistance (Fig.
1.2c). Low genetic diversity was at the basis of the Vancouver outbreak of Cryptococcus gattii (Kidd et al. 2004), and the Oregon outbreak concerned a single highly virulent genotype (Byrnes et al. 2010) (Chap. 12). Emerging fungal infections in humans and other vertebrates often take us by surprise. Bat white-nose disease suddenly killed thousands of bats in the USA and was caused by a species that was known in Europe and to which European bats were resistant (Chap. 13). The fungus appeared extremely successful in susceptible American bat populations, likely owing to physiological and behavioral differences between new- and old-world bats. The causative agent, Pseudogymnoascus destructans, was described only in 2009 (Gargas et al. 2009) and the fungus seemed to appear out of the blue. Several of the species that cause large epidemics or epizootics have only recently been described; first cases were often recorded only a few decades ago. One of the examples is Aspergillus felis causing infections in cats in Australia (Barrs et al. 2007) (Chap. 15). One might suppose that this is a fallacy due to insufficient awareness and inadequate diagnostics in the past, but in large collections of, for example, archived Sporothrix material, the recent outbreak fungus Sporothrix brasiliensis was not detected (Rodrigues et al. 2014). The frog disease agent Batrachochytrium dendrobatidis was described only in 1999 (Longcore et al. 1999) with its newly introduced counterpart B. salamandrivorans killing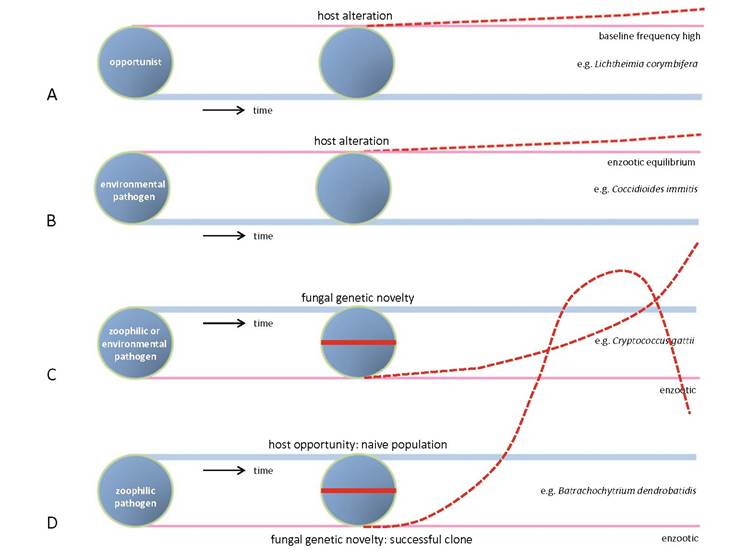
Fig. 1.2 Predicted emergence of species.
(a) Opportunist with rather high frequency, increase relative to growth of susceptible populations; (b) environmental pathogen, relatively frequent in endemic area, in equilibrium with its habitat, increase relative to growth of susceptible populations; (c) pathogen developing novelty by which it increases the number of susceptible hosts; (d) zoophilic pathogen developing novelty, confronted with naive host populations which leads to epidemic expansion, expected decrease when susceptible hosts are no longer availablesalamanders in 2013 (Martel et al. 2013) (Chap. 14). These pathogens have likely been present in low abundance and were undetected until the encounter of a virulent genotype and susceptible host found a match, which was thought to have occurred due to trade-facilitated intercontinental movements. Both species have low genetic diversity suggesting clonal expansion (Morehouse et al. 2003; Fisher et al. 2012). Over a larger sampling area, there is genetic diversity, but local frog mortality is caused by single clones (Morgan et al. 2007). Fastest population expansion is predicted to occur when virulent genotypes of pathogens match with a novel window of opportunity (Fig. 1.2). An extra window of opportunity for frog disease has been suggested to be climate change, weakening frog populations (Clare et al. 2016). The combined changes in fungus and host allow asymptotic expansion of the pathogen, finally leading to decline because susceptible hosts have become rare (Fig. 1.2d). This may account for the wavelike pattern of lethargic crab disease along the Brazilian coast (Avila et al. 2012) (Chap. 11).
The emergence of sporotrichosis is still largely unexplained. Cat-transmitted Sporothrix brasiliensis shows epizootic and epidemic expansion in Southeast Brazil since 1990 (Ortiz Sanchotene et al. 2015) (Chap. 10). The species is one of two near- clonal entities that have evolved from Sporothrix schenckii (Rodrigues et al. 2014; Moussa et al. 2017). In the ancestral species, both cat and plant transmission are documented. The clones have specialized in either one of these modes of transmission: both are traumatically inoculated into the skin, but S. brasiliensis enters via cat scratches, whereas S. globosa infection stems from trauma caused by sharp plant debris or thorns. Sporothrix globosa infection is seasonal (Yu et al. 2013), whereas the more virulent S. brasiliensis replicates asymptotically to become overabundant, outcompeting the ancestral species, S. schenckii (de Araujo et al. 2015). The remarkable difference in transmission mode, with large consequences for evolutionary success and public health is observed at a very small phylogenetic distance (Moussa et al. 2017).
1.5