Recent Outbreaks and Epizootics
Candida albicans, commonly causing mucocutaneous infections in humans, is transmitted directly, from patient to patient, e.g., from mother to child. The host is niche as well as the reservoir of the fungus.
The species has an advantage of infecting the vertebrate host and can thus be interpreted as a zoophilic pathogen (Fig. 1.1). Carriage by the healthy host is mostly non-symptomatic, the fungus then maintaining as a commensal (Achkar and Fries 2010) and every new host may be contagious. With only slight impairment of innate or acquired immune systems of the host, C. albicans infections become symptomatic. With host-to-host transmission, there is no common source of contamination, as opposed to sapronoses. In the susceptible population of hospitalized patients, other Saccharomycetales are emerging, such as the fluconazole-resistant Candida auris (Chowdhary et al. 2016). The species has now been reported globally from bloodstream infections in severely compromised patients. It often occurs in the form of local outbreaks (Schelenz et al. 2016). Its mode of infection is as yet unknown, but host-to-host transmission seems an option.Emergomyces africanus is a novel human pathogen involved in an outbreak of disseminated disease in HIV-positive individuals in South Africa (Kenyon et al. 2014; Dukik et al. 2017). Its origin is unknown; probably the fungus is an environmental pathogen and might occur in animal populations. The genus Emergomyces contains a number of species which since recently were repeatedly involved in human infection, while related species reside fairly commonly in the lungs of small rodents (e.g., Emmonsia crescens) (Borman et al. 2009, Hubalek et al. 1998) (Chap. 7), infection of humans being coincidental (Dot et al. 2009). Screening of wild animals is required to understand, and to possibly predict, host jumps to humans.
Humans are likely non-optimal hosts in most species, and relations between animal hosts may be quite complicated (Vilela et al. 2016). Jiang et al. (2018) noted that small temperature differences of dimorphic switch in Emergomyces and Blastomyces species may explain predilections for different host animals and thus may be a driver of new directions in the evolution of these fungi. Ecological fitting (Araujo et al. 2015) via the sloppy fitness space of the not-yet-suitable host might be the optimal model to describe the sympatric evolutionary processes in these fungi.Most Emergomyces species and other members of Ajellomycetaceae such as Paracoccidioides (Chap. 6) are environmental pathogens with a double life cycle (Fig. 1.3), i.e., living permanently in the environment but having a certain advantage (increased fitness) if an animal host is used in any stage of the life cycle. The host may be regarded as a vehicle, which enhances dispersal, optimizing the distribution of the fungus in the environmental niche. For example, Histoplasma capsulatum is found on sheltered animal droppings, e.g., bat feces in caves or bird roosting sites (Rocha-Silva et al. 2014). Often the resident animals are asymptomatically colonized (Naiff et al. 1996), return to the cave, and ultimately die there (Chap. 5). Thus, there is no efficient vehicle of dispersal, but susceptible visitors of the cave after contamination and infection are likely to die elsewhere, enabling further dispersal of the pathogen. The fungus thus has a strategy to reach optimal fitness. A common-source outbreak is concerned with the number of infections corresponding with the number of susceptible individuals entering the contaminated site; thus usually no epidemic or
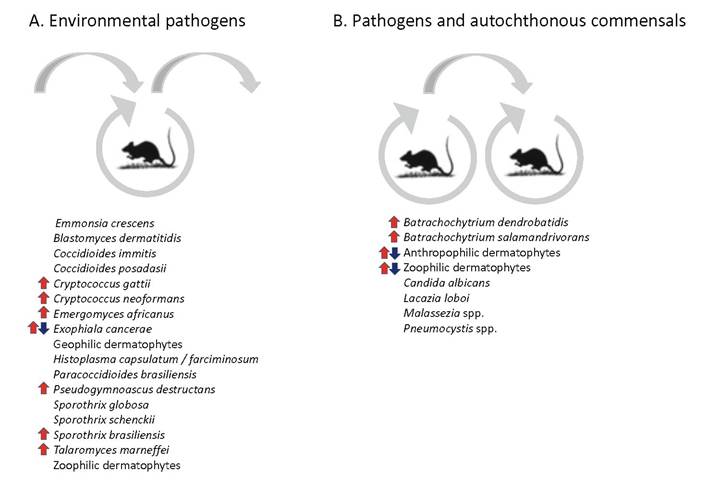
Fig. 1.3 Current approximate changes in frequency in (a) environmental pathogens and (b) zoophilic pathogens. Large Exophiala cancerae zoonosis has disappeared; dermatophyte floras on humans are changing due to socioeconomic changes
epizootic expansion is noted. Hosts are not contagious, but are infected by environmental propagules; the infections are sapronoses.
In the main common-source, sapronotic outbreaks of environmental pathogens such as Histoplasma, the role of the susceptible host in fungal expansion is rather insignificant. The size of the inoculum is proportional to the presence of the fungus in its environmental niche. Outbreaks of Coccidioides immitis are related to weather conditions and subsequent dust storms that lead to massive inhalation of propagules (Valdivia et al. 2006) (Chap. 4). Over time, the fungal prevalence remains more or less the same, though with fluctuations. Coccidioides is hypothesized to form hyphae from cells located in host tissue after its death, to colonize a new environmental site (Lewis et al. 2015). The infection is controlled primarily by acquired cellular immunity and therefore tends to expand with an increasing number of AIDS patients (Rempe et al. 2007).
Talaromyces marneffei is an unrelated environmental pathogen. The species occurs in soil and has a reservoir in large rodents in Southeast Asia but found a susceptible new host due to the human AIDS pandemic. Dormancy inside living pulmonary tissue of the animal requires resistance to innate immunity and control by acquired cellular immunity (Cooper and Vanittanakom 2008), and hence impairment of T-cell function provides an obvious portal for dissemination. Also, this species is overwhelmingly clonal (Fisher et al. 2005) despite local sexuality. Populations are geographically substructured with likely adaptation to different host species (Henk et al. 2012). The environmental habitat of the agent of bat white-nose syndrome (WNS), Pseudogymnoascus destructans causing bat decline in northeastern USA (Blehert et al. 2009), is still unknown (Chap. 13) (Rajkumar et al. 2011). The virulent clone causing the bat near extinction presumably already pre-existed in Europe, where bats had found a balance toward infection without causing significant problems. However, when reaching susceptible bat populations in the USA, the fungus was able to expand exponentially (Leopardi et al.
2015).Recent outbreaks have also been caused by dermatophytes, which are keratinophilic fungi with a gradual adaptation to living tissues (Chap. 3). Anthropophilic dermatophytes are highly specialized species with niche and reservoir on human skin. Transmission takes place from human-to-human, or via propagules which can survive in the environment but do not form assimilative thalli, comparable to Batrachochytrium. Zoophilic dermatophytes are adapted to animal skin, with different level of host-specificity. Trichophyton equinum is isolated from horses only, whereas Microsporum canis can be detected in a wide range of animals (and sometimes in humans). Zoophilic dermatophytes are mostly carried in animal fur, which is often in close contact with soil and plant material of the host's burrow. Furred animal thus may be infected directly from their nest-mates but also from their environment. Carriage in the fur is often asymptomatic, infection only taking place in susceptible nest-mates, e.g., juveniles, or when a non-suitable host is coincidentally infected, such as a predator or a human. Geophilic dermatophytes have a double life cycle with elaborate sexual phases that are produced in soil. Sexuality in anthropophilic dermatophytes is not known to exist or concerns somatic cell fusion at most. A gradual loss of sexuality in dermatophytes with advancing adaptation to mammalian hosts has been observed (de Hoog et al. 2016). Clonal reproduction is often prevalent (Graser et al. 2006), which is underlined by the fact that potential mating partners having either high mobility group (HMG) or α-box transcription factors can be phenotypically different (Symoens et al. 2013).
1.6