Fluid and Electrolyte Balance
Packed Cell Volume and Total Plasma Protein
PCV and total plasma protein (TPP) concentration are simple and convenient indicators of acute fluid alterations. Changes in the plasma volume generally are reflected by changes in the PCV and the TPP concentration.
In dehydrated humans, changes in the PCV are believed to be the more reliable guide to changes in plasma volume because substantial protein fluxes into and out of the circulation have been shown to occur. However, in most animal species the range of normal for the PCV is much wider than for the TPP concentration. This is particularly true of horses, in which excitement, pain, or catecholamine release can produce variable mobilization of splenic erythrocytes, making it difficult to obtain a truly resting PCV. For these reasons, precise quantitative estimation of a change in plasma volume using these parameters is more complex and less reliable in large animal species. As plasma volume increases or decreases, the change in the PCV is always less than the change in the TPP concentration. However, a large disparity in the changes in the PCV and the TPP concentration in a patient with a history of loss of sodium-containing fluid and clinical evidence of reduced effective circulating fluid volume suggests blood or protein loss. Marked increases in the PCV with a normal-to-low TPP concentration frequently are encountered in animals with acute protein-losing enteropathies such as salmonellosis or equine toxic enteritis, or enterocolitis. In horses undergoing treatment for diarrhea, the excessive administration and retention of sodium-containing fluids is a key factor in the development of edema and hypoproteinemia. Blood loss generally results in a decrease in both PCV and TPP concentration with a lag time of up to 24 to 48 hours.Serum Sodium
The serum sodium concentration is a function of the exchangeable cation content (i.e., the exchangeable sodium [Na] in the extracellular fluid [ECF] volume plus the exchangeable
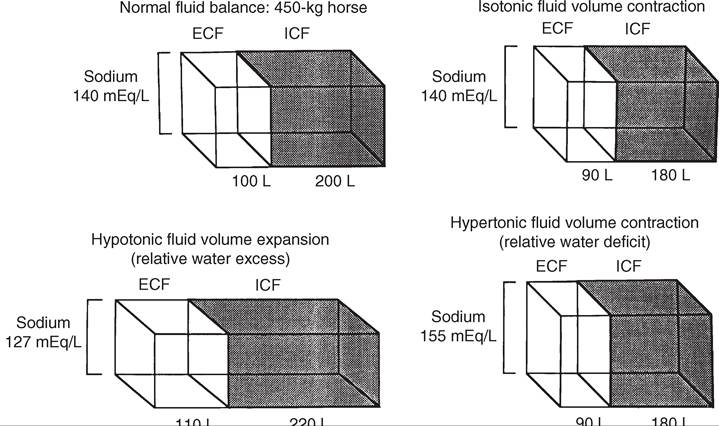
FIG.
22.1 Compartmental distribution of fluid between the extracellular fluid (ECF) volume and the intracellular fluid (ICF) volume in a 450-kg horse with normal fluid balance, isotonic fluid volume contraction, hypotonic fluid volume expansion, and hypertonic fluid volume contraction. (Modified from Kaneko JJ, Harvey JW, Bruss ML, editors: Clinical biochemistry of domestic animals, ed 6, New York, 2008, Elsevier Academic Press.)potassium [K] in the intracellular fluid [ICF] volume relative to total body water). The ECF sodium content determines the ECF volume, whereas the ICF volume is a function of ICF potassium content.
Changes in the sodium concentration reflect the net changes in this relationship and often do not represent accurately the changes in sodium balance. Changes in water balance are thus primarily responsible for changes in the serum sodium concentration, and one should always consider the hydration status of the patient for sodium concentration interpretation. Hyponatremia is an indication of a relative water excess, whereas hypernatremia is an indication of a relative water deficit.
Dehydration is defined as a loss of body water (fluid volume contraction). It occurs in a variety of clinical circumstances. The serum sodium concentration provides a means of categorizing dehydration in a physiologically meaningful way. Hypertonic dehydration, which occurs when water losses exceed the losses of sodium and potassium, is indicated by hypernatremia (free water loss or hypotonic fluid losses). The response of horses to feed and water deprivation is an example of this form of dehydration. Isotonic dehydration occurs with a balanced loss of water and electrolytes—that is, approximately 140 to 150 mEq of sodium plus potassium (Na + K) for each liter of water lost. Because the relative water balance has not changed, the serum sodium concentration remains unchanged despite the accumulation of what may have been a substantial body sodium deficit. The early stages of acute diarrhea and the dehydration of heavily sweating endurance horses are examples of isotonic dehydration.
Hypotonic dehydration occurs when the losses of exchangeable cations (Na + K) exceed the net change in water balance; this condition is indicated by hyponatremia. Hypotonic dehydration often is seen in animals with subacute or chronic diarrhea that develop substantial water and electrolyte deficits but then replace part of the water deficit through water consumption. Fig. 22.1 shows the compartmental distribution of fluid between the ECF volume and ICF volume in four situations.HYPONATREMIA. Hyponatremia results from retention of free water or excess loss of sodium. It is often but not invariably associated with conditions that cause sodium depletion such as vomiting, diarrhea, excessive sweat losses, and adrenal insufficiency. The fluid losses in these conditions are most often hypotonic or isotonic, and initial fluid and electrolyte deficits do not result in hyponatremia until water intake, renal water retention, or both disturb the balance between the remaining exchangeable cations and the total body water.
The accumulation of sodium-containing fluid in body cavities or the gut lumen caused by ascites, peritonitis, or rupture of the bladder or by displacement, torsion, or volvulus of the gut is referred to as a third-space problem. When such accumulations develop rapidly, the plasma volume is reduced and the serum sodium concentration subsequently may decrease as compensating renal responses cause water retention. Rupture of the bladder in neonatal foals is associated with marked hyponatremia and hypochloremia. As fluid intake continues and dilute urine accumulates in the abdomen, sodium, chloride, and other ions are drawn from the rest of the ECF into this accumulating fluid. No sodium or chloride has been lost from the body, and the observed decreases in the electrolyte concentration are caused by changes in the relative water balance (“water follows solute”). The neurologic signs seen in these foals are largely caused by the effects on the central nervous system of the rapidly developing and marked hypotonic hyponatremia.
Progressively severe neurologic disturbances may be seen as the serum sodium concentration falls below 115 mEq/L and then below 100 mEq/L. The severity of the neurologic abnormalities is a function of both the rate at which hyponatremia develops and the absolute degree of hyponatremia. Neurologic disturbances can occur iatrogenically if excessive amounts of free water (usually given as 5% dextrose) are administered to patients with altered renal function.Mastitis results in an increased loss of sodium in the milk, and a low-grade mastitis problem in a dairy herd on a marginal dietary salt intake may result in sodium depletion and medullary washout. Decreased milk production, polyuria, hyposthenuria, and a low urine sodium level may be noted, although the serum sodium concentration may remain within the lower range of normal.
The most common causes of hyponatremia are listed in Box 22.1. Marked hyperlipidemia or hyperproteinemia produces a falsely low sodium concentration value because lipid or protein occupies a significant volume in the serum or plasma sample and because sodium is present only in the aqueous phase. This potential cause of hyponatremia is indicated by an increase in
■ BOX 22.1
■ BOX 22.2
Causes of Hyponatremia
Common Causes
Relative water excess
Loss of sodium-containing fluid (decreased effective circulating volume)
Diarrhea
Excessive sweating
Blood loss
Fluid drainage
High-volume gastric reflux
High-volume pleural drainage Adrenal insufficiency
Sequestration of fluid (third-space problems)
Peritonitis
Ascites
Pleuritis
Ruptured bladder
Torsion or volvulus of the gut
Excessive administration of 5% dextrose to patient with renal disease
False hyponatremia Hyperlipidemia Hyperproteinemia Hyperglycemia
Uncommon Causes
Water retention with normal effective circulating volume Psychogenic polydipsia
Renal disease
Inappropriate antidiuretic hormone secretion Use of diuretics
the osmolar gap between measured and calculated osmolality.
The use of direct ion-selective electrodes (direct potentiometry; all modern blood gas machines; and modern multianalyzers of big laboratories) for electrolyte determinations avoids this potential cause of a falsely low sodium concentration value.Marked hyperglycemia causes a reduction in the measured serum sodium concentration of approximately 1.6 mEq/L for every 100 mg/dL increase in the glucose concentration. Increases in the plasma glucose concentration generate osmotic forces that result in the movement of cellular water into the ECF, diluting the plasma sodium concentration.
HYPERNATREMIA. Hypernatremia can develop if water is lost in excess of sodium or if sodium is ingested in excess of water. Hypernatremia can occur in the initial stages of diarrhea, vomiting, or renal disease if water loss exceeds electrolyte loss (Box 22.2). When water losses are replaced by increased water consumption, enhanced renal water retention, or both, the serum sodium concentration decreases. Food and water deprivation in normal horses and cattle is associated with substantial reduction of renal and fecal output, but continued cutaneous and respiratory insensible water loss may result in hypernatremia (primary water loss). Hypernatremia may occur transiently as a result of sodium excess after administration of hypertonic saline or hyperosmolar sodium bicarbonate if water intake is restricted or impaired. Hypernatremia has been reported in calves fed an inappropriately mixed oral electrolyte replacement solution as their only fluid intake.13 The hypernatremia observed with salt poisoning in cattle and swine is the result of water restriction in animals that have been maintained on a high-salt intake.
Serum Potassium
Potassium is the major intracellular cation. Between 60% and 75% of total body potassium is found within muscle cells and
Causes of Hypernatremia
Common Causes
Pure water losses
Panting
Water deprivation
Sodium excess (water restriction)
Salt poisoning
Feeding only electrolytes, no free water
Uncommon Causes
Water loss exceeds electrolyte loss
Vomiting
Diarrhea
Burns
Intrinsic renal disease
Diuretics
Diabetes insipidus
Central
Nephrogenic
Hypertonic saline or sodium bicarbonate administration Mineralocorticoid excess
in bones.
Only 5% is in the ECF and is not always a reflection of total body potassium. The serum potassium concentration is influenced by factors that alter internal balance (the distribution of potassium between the ECF and the ICF) and those that change external balance (potassium intake and output). Potassium is higher in foals compared with adult horses. Generally, herbivores have a net positive intake in potassium, which is usually carefully regulated by the renal system. Serum potassium concentrations correlate relatively well with total body potassium content in cattle but not in horses. Changes in the serum potassium concentration occur in a wide variety of clinical circumstances and have profound neuromuscular effects that are largely the result of changes in cell membrane potential. The responses to dehydration and acid-base imbalance often complicate the evaluation of the potassium concentration. For example, calves with acute diarrhea often develop body potassium depletion because of excessive losses and inadequate intake, but the serum potassium concentration of these animals is usually normal to increased as the result of renal shutdown and the metabolic acidosis induced by dehydration, sodium depletion, and hypovolemia. Hypokalemia may become evident only as other fluid and electrolyte losses are replaced.Measuring the erythrocyte potassium concentration is relatively easy and has been suggested as an aid in assessing the need for potassium supplementation in racehorses with recurrent muscle disease or in alert downer cows early postpartum. However, experimental studies in horses indicate that the erythrocyte potassium concentration does not always accurately reflect potassium deficits.
HYPOKALEMIA. Hypokalemia increases resting membrane potential, resulting in muscle weakness, impaired urine concentration ability, and arrhythmias. Hypokalemia may result from depletion of the body's potassium stores or from a redistribution of potassium from the ECF into the ICF space (Box 22.3). Hypokalemia is most commonly seen with altered intake and absorption and with excessive potassium losses from the gastrointestinal tract caused by vagal indigestion, torsion of the abomasum, ileus, or diarrhea. Excessive renal loss may result from mineralocorticoid excess, certain diuretics, or altered renal function, as reported in horses with renal tubular acidosis. Marked hypokalemia develops when reduced dietary intake
■ BOX 22.3
■ BOX 22.4
Causes of Hypokalemia
Common Causes
Altered External Balancea
Vomiting
Vagal indigestion with internal vomiting
Diarrhea
Third-space problems (gut or abomasal torsion or volvulus; peritonitis)
Excessive sweat losses
Dietary deficiency
Prolonged anorexia
Altered Internal Balance
Metabolic alkalosis
Uncommon Causes
Altered External Balance
Mineralocorticoid excess
Use of corticosteroids with mineralocorticoid activity Diuretics
Renal tubular acidosis
Postobstruction diuresis
Altered Internal Balance
Excessively rapid sodium bicarbonate administration
Insulin and/or glucose administration Catecholamine administration or endogenous release
aExternal balance refers to the relative changes in potassium intake and output; internal balance refers to the distribution of potassium between the extracellular and intracellular fluid compartments.
caused by anorexia is associated with excessive potassium losses (e.g., causing muscle paresis in dairy cows early postpartum). A hypokalemia syndrome has been observed in cattle,14 mainly in postpartum lactating cows, but also in younger animals. The exact causes have not been completely elucidated except for isoflupredone administration. The syndrome is characterized by muscle weakness, severe depression, recumbency, tachycardia, cardiac arrhythmia, and abnormal neck posture. The lowest reported serum potassium concentration levels are between 2.2 and 2.5 mmol/L.
Hypokalemia without potassium depletion results from the movement of extracellular potassium to the intracellular space. This form of hypokalemia occurs in response to an acute alkalosis and the administration of insulin or glucose. Overzealous and rapid administration of sodium bicarbonate can produce an alkalosis with a profound and rapidly developing hypokalemia. Animals with moderate potassium deficits that are vigorously treated with sodium bicarbonate to correct a coexisting mild metabolic acidosis may be particularly prone to this problem. The initial response to catecholamine administration is a modest, transient increase in potassium caused by α-adrenergic stimulation, which is often followed by hypokalemia caused by β-adrenergic receptor responses.
Pseudohypokalemia is seen in severe lipemia with potassium measured with indirect potentiometry (e.g., older automated profile chemistry analyzers).
HYPERKALEMIA. Hyperkalemia may develop in vitro as a result of hemolysis or leakage of erythrocyte potassium after storage of whole blood (Box 22.4). The release of potassium from leukocytes or platelets into the serum after clot formation is a potential cause of hyperkalemia if marked leukocytosis or thrombocytosis is present. Hyperkalemia also results from renal potassium retention in Addison disease, acute renal failure (ARF), and renal shutdown. A number of factors contribute
Causes of Hyperkalemia
Common Causes
False hyperkalemia
In vitro hemolysis
Prolonged storage of blood (over 6 hours) without separation of serum or plasma
Altered external balance
Hypovolemia with renal shutdown Altered internal balance
Metabolic acidosis
Vigorous exercise
Uroperitoneum
Uncommon Causes
False hyperkalemia
Markedly elevated leukocyte or platelet count Altered internal balance
Hyperkalemic periodic paralysis in Quarter Horses Diabetes mellitus
Tissue necrosis
Renal disease
Addison disease
to the movement of intracellular potassium into the ECF, resulting in hyperkalemia. Hyperkalemia is often associated with metabolic acidosis, particularly when the acidosis results from volume depletion and is complicated by renal shutdown. Hyperkalemia has been reported in animals with massive muscle necrosis, but neither hyperkalemia nor metabolic acidosis is a common feature in horses with exertional rhabdomyolysis. Vigorous short-term exercise of horses at high intensity results in a marked but transient hyperkalemia (9 to 10 mEq/L) that may be associated with the profound lactic acidosis seen with anaerobic workloads.15 Potassium returns to normal within minutes, and often a modest hypokalemia occurs later in the recovery period. Episodic hyperkalemia and muscular weakness are associated with the condition of HYPP.16 HYPP is inherited as an autosomal-dominant trait in horses, with a specific Quarter Horse lineage having a sodium channel defect17 (see Chapter 42 for a more complete discussion of this disorder). The disease is the result of a single DNA base pair substitution that leads to the production of an abnormal voltage-regulated sodium channel at the cell membrane.18 Sudden marked increases in the serum potassium concentration, up to 8 to 9 mEq/L, are the result of transcellular movement of potassium and are associated with profound electrocardiographic abnormalities and fluid shifts.
In drug-induced hyperkalemia, trimethoprim has been associated with hyperkalemia by inhibiting normal sodium reabsorption in the cortical renal collecting duct.
Serum Chloride
Chloride is a major ECF anion and important with sodium for osmolality and acid-base balance. Together with sodium it is the main component of the strong ion difference (SID). Alterations in the chloride concentration are usually associated with nearly proportional changes in the sodium concentration as the result of changes in relative water balance. In addition, the chloride concentration tends to vary inversely with the bicarbonate concentration; therefore when disproportionate changes in the chloride concentration relative to sodium occur, significant acid-base imbalances should be anticipated. Disproportionate increases in chloride are associated with a normal-to-low anion gap hyperchloremic metabolic acidosis, but they also are seen as a result of the compensating responses for a primary
■ BOX 22.5
■ BOX 22.6
Causes of Hyperchloremia
With Proportional Increase in Sodium
Common Causes
Relative water deficit
Panting Water deprivation Salt poisoning
Uncommon Causes
Vomiting
Diarrhea
Burns
Intrinsic renal disease
Diuretics
Diabetes insipidus
Central
Nephrogenic Hypertonic saline administration Mineralocorticoid excess
Without Proportional Increase in Sodium Common Causes
Hyperchloremic metabolic acidosis
Renal tubular acidosis
Uncommon Cause
Compensation for respiratory alkalosis
respiratory alkalosis (Box 22.5). A striking hyperchloremic metabolic acidosis has been reported in horses with renal tubular acidosis.19,20 Iatrogenic hyperchloremic acidosis can be induced by overzealous NaCl fluid treatment.21
Disproportionate decreases in chloride relative to sodium are characteristically seen in metabolic alkalosis but also may be seen as part of the compensating response for chronic primary respiratory acidosis (Box 22.6). Hypochloremic metabolic alkalosis is a common feature in many digestive disorders of ruminants and is caused by loss of chloride-rich fluids or sequestration of such fluids in the abomasum and forestomachs. It is also seen in horses with diarrhea as chloride is absorbed in the ileum and colon. Horses treated with diuretics (e.g., furosemide-induced metabolic alkalosis) may show hypochloremia.22 Heavy sweating in horses is associated with hypochloremic metabolic alkalosis.
Osmolality
ECF osmolality is maintained around 300 mOsm (isotone). Measurement of the serum osmolality provides an indication of relative water balance in much the same way the serum sodium concentration does. In most circumstances these parameters are closely correlated. Comparing the measured osmolality with the calculated osmolality, as determined from the measured concentrations of the major solutes in serum (sodium, chloride, glucose, and urea), provides a means of determining if the serum water content deviates widely from normal or if foreign, low-molecular-weight substances are present in the blood. The difference between the measured osmolality and the calculated osmolality is called the osmolar gap. Decreases or increases in the osmolar gap could indicate laboratory error, but increases of >10 mOsm/kg generally are the result of either a decrease in serum water content (caused by hyperlipidemia or hyperproteinemia) or the presence of abnormally high concentrations of low-molecular-weight substances in the serum. These substances can include a variety of exogenous and potentially toxic compounds such
Causes of Hypochloremia
With Proportional Decrease in Sodium
Common Causes
Relative water excess
Diarrhea Excessive sweating Blood loss Fluid drainage
High-volume gastric reflux
High-volume pleural drainage
Sequestration of fluid (third-space problems) Peritonitis
Ascites
Ruptured bladder Renal disease
False hypochloremia Hyperlipidemia Hyperproteinemia Hyperglycemia
Uncommon Causes
Psychogenic polydipsia
Inappropriate antidiuretic hormone secretion Adrenal insufficiency
Without Proportional Decrease in Sodium
Common Causes
Metabolic alkalosis
Exhaustive disease syndrome Abomasal torsion
Vagal indigestion with internal vomiting Response to furosemide in horses
Uncommon Cause
Compensation for respiratory acidosis
as mannitol, ethanol, methanol, propylene glycol, ethylene glycol, isopropanol, ethyl ether, acetone, trichloroethane, and paraldehyde. All hypernatremic animals are hyperosmolal. Hypo-osmolality is always associated with hyponatremia, but not all cases with hyponatremia are hypo-osmolal (e.g., concurrent hyperglycemia).
Serum Calcium
Calcium plays a vital role in many of life's processes including maintenance of neuromuscular excitability, permeability of cell membranes, conduction of nerve impulses, muscle contraction, and blood clotting. For these reasons the serum calcium concentration or, more correctly, the ionized calcium concentration is normally maintained within a relatively narrow range, despite wide variation in intake and output. Calcium metabolism is regulated by dietary factors, vitamin D and its active metabolites, and the hormones parathormone and calcitonin. Compared with ruminants in horses, a relatively larger percentage of calcium is excreted in the urine. The serum calcium concentration is maintained by adjusting intestinal absorption, renal excretion, and mobilization of available calcium from the large stores in bone. Calcium exists in the serum in three forms: ionized calcium, complexed calcium, and protein-bound calcium. Ionized calcium, which normally constitutes 50% to 60% of the total calcium, is the physiologically active form of calcium. Protein binding can cause confusion (protein-bound calcium normally constitutes 40% to 50% of the total calcium, and most of it is bound to albumin). Hyperalbuminemia may result in a modest hypercalcemia, whereas hypoproteinemia, especially hypoalbuminemia, regularly results in a moderate hypocalcemia. The ionized calcium concentration, usually measured with ion selective electrodes, generally remains within normal limits, despite increases or decreases in total calcium associated with the change in protein concentration. The acid-base balance has additional influence on the amount of ionized and proteinbound calcium. Alkalosis reduces ionized calcium and increases protein binding, whereas acidosis produces the opposite effect. Ion-specific electrodes are available for determining the ionized calcium level, which can be useful if blood samples are handled appropriately. Most diagnostic laboratories provide the total serum calcium value, which is composed of ionized, complexed, and protein-bound calcium.
HYPOCALCEMIA. Large increases or decreases in the serum calcium concentration generally are the result of a failure in the normal mechanisms of calcium homeostasis rather than a reflection of absolute calcium deficits or calcium-phosphorus imbalances. Hypocalcemia occurs with some frequency in domestic animals, particularly in high-producing dairy cattle at the onset of lactation. In cattle the serum calcium concentration normally decreases to sites, thus reducing the incidence of milk fever in cows at high risk. Grass tetany is associated with marked hypomagnesemia and modest hypocalcemia, whereas the inorganic phosphorus level remains within the normal range.
Systemic diseases resulting in anorexia (e.g., traumatic reticuloperitonitis, ketosis, and displaced abomasum) or acute toxemic conditions (e.g., coliform mastitis, septicemia,
■ BOX 22.7
Causes of Hypocalcemia
Common Causes
Parturient paresis (milk fever)
Grass tetany
Hypoalbuminemia (decreased total calcium; ionized calcium may remain unchanged)
Fat necrosis
Lactation tetany
Transport tetany
Synchronous diaphragmatic flutter
Blister beetle toxicosis (cantharidin)
Acute renal failure
Anorexia in lactating cows
Colic in horses
Uncommon Causes
Acute toxemia and associated anorexia in lactating dairy cows
Hypoparathyroidism
Exertional rhabdomyolysis
Malignant hyperthermia
Pancreatic disease
Oxalate toxicity
Tetracycline administration
Furosemide administration
Alkalosis induced by excessive bicarbonate administration or aspiration pneumonia) that produce anorexia in lactating cattle frequently result in hypocalcemia. Hypocalcemia is also seen in sheep on marginal rations if stressed by inclement weather or when being moved and during late pregnancy as well. Hypocalcemia is seen in cattle with fat necrosis, presumably as the result of incorporation of calcium with the fat as a form of soap. Horses sweating or with exhaustive disease syndrome or transit tetany often develop decreases in ionized calcium with resultant muscle cramps and synchronous diaphragmatic flutter, which generally respond to intravenous calcium and electrolyte fluid administration. Horses, cattle, and sheep usually respond initially to acute renal tubular damage with mild hypocalcemia and hyperphosphatemia. Idiopathic hypocalcemia, which may exist in foals 4 to 5 days old, has poor prognosis, and a cause is not established.23 Transport tetany and lactation tetany of horses are associated with hypocalcemia. In lactation tetany of mares, this is correlated with high milk production.
HYPERCALCEMIA. Marked hypercalcemia, with a serum calcium level ranging from 14 to 20 mg/dL (3.5 to 5 mmol/L), and modest hypophosphatemia are frequently observed in horses with chronic renal failure (CRF) that are fed a high- calcium diet such as alfalfa hay (Box 22.8). In these horses, blood samples collected in standard EDTA tubes may actually clot. This occurs because the serum calcium concentration is so high that there is insufficient EDTA to bind all the calcium, and some free calcium is available to complete the clotting process. Vitamin D intoxication can develop as a result of excessive dietary supplementation or the ingestion of certain plants such as Cestrum diurnum (day blooming jasmine), Solanum malacoxylon, Solanum sodomaeum, and Trisetum flavescens, which contain toxic quantities of vitamin D analogs. Primary hyperparathyroidism is exceedingly rare in large animals, but pseudohyperparathyroidism with hypercalcemia occasionally can develop in animals with tumors that produce protein substances with parathormone-like biologic activity. This has been reported in a few horses with lymphosarcoma, squamous cell carcinoma, adrenocortical carcinoma, multiple myeloma, and ameloblastoma. Hypercalcemia has also been reported in idiopathic systemic granulomatous disease and some foals with neonatal asphyxia.24
Serum Phosphorus
Phosphorus is found primarily in the skeleton and teeth in close association with calcium in the intricate and dynamic crystalline structure of bone. Intracellularly, phosphate plays an essential role in the degradation and synthesis of many compounds. Also, in the forms of adenosine triphosphate (ATP), adenosine diphosphate (ADP), and adenosine monophosphate
■ BOX 22.8
Causes of Hypercalcemia
Common Causes
Chronic renal failure in horses
Hypervitaminosis D
Excessive dietary supplements
Plant intoxication
Cestrum diurnum (day blooming jasmine)
Solanum malacoxylon
Excessive or too rapid intravenous administration of calcium
Uncommon Causes
Neoplasia (pseudohyperparathyroidism) Lymphosarcoma
Gastric squamous cell carcinoma
Hyperparathyroidism
■ BOX 22.9
■ BOX 22.10
Causes of Hypophosphatemia
Common Causes
Chronic renal failure in horses
Parturient paresis in cattle
Postparturient hemoglobinuria
Uncommon Causes
Brassica toxicity
Inadequate dietary intake
Starvation or chronic wasting diseases
Hyperparathyroidism, pseudohyperparathyroidism
Causes of Hyperphosphatemia
Common Causes
Acute renal failure
Nutritional secondary hyperparathyroidism (excess phosphate intake)
Endurance exercise in horses
Higher normal range in neonates
Uncommon Causes
Acute rhabdomyolysis
Vitamin D toxicity
(AMP), it is the primary form of energy storage and transfer required for almost all of life's processes. Phosphorus in the ECF exists primarily in the organic and inorganic form, specifically as the “buffer” pair H2PO-3 and HPO-4, and plays a role in acid-base balance. Like calcium, phosphorus is regulated by dietary factors, the active metabolites of vitamin D, and the hormones parathormone and calcitonin. Imbalances of calcium and phosphorus or the presence of compounds that bind these substances in the gut can produce serious imbalances that are not always evident on analysis of serum samples. Measurement of urinary output or creatinine clearance ratios for calcium and phosphate are simple and helpful procedures. They provide an indication of an imbalance while more definitive procedures such as ration analysis are contemplated.
HypophosphATEMIA. Serum phosphorus concentrations are not always an accurate guide to phosphate balance, but dietary deficiencies of phosphorus are frequently manifested by hypophosphatemia. Hypophosphatemia is a common feature in cattle with parturient paresis (see also Chapter 41) and horses with CRF (Box 22.9). It has been reported in animals with experimental oxalate toxicity, in chronic wasting states, or with starvation. Postparturient hemoglobinuria is a disorder of cattle, primarily lactating dairy cattle, that often is associated with diets low in phosphorus. Although marked hypophosphatemia is often reported, it is not an invariable feature of this disorder. Hypophosphatemia has also been seen in cattle on acidogenic diets as a consequence of increased excretion and can also be observed due to compartmental phosphorus shifts as in hypertonic dextrose administration.25 Hypophosphatemia in horses has also been observed in renal failure, sepsis, malignancies, hyperinsulinism, metabolic alkalosis, and parenteral nutrition.24
HYPERPHOSPHATEMIA. Age -related differences exist in the normal range of serum phosphorus concentration. Young animals have much higher values than adults, with values for neonates commonly up to 7 to 9 mg/dL (2.3 to 3.0 mmol/L). The serum phosphate concentration declines progressively with age. Hyperphosphatemia is seen in animals with vitamin D toxicity, transiently in horses after long-distance endurance rides, and initially in horses with ARF (Box 22.10). It has also been observed in hypothyroidism, marked tissue necrosis (e.g., rhabdomyolysis, intestinal infarction, tumor lysis syndrome, hemolysis), hypoparathyroidism, and in foals treated with phosphate-containing enemas. In ruminants, hyperphosphatemia is more commonly seen in dehydration and ensuing hemoconcentration as in abomasal displacement and volvulus, where it has been shown to be closely associated with dehydration.26
Serum Magnesium
Between 20% and 30% of magnesium is bound to mostly albumin, 10% is bound to weak acids, and 60% is free (ionized).27 Free magnesium can be measured with ion-selective electrodes.
■ BOX 22.11
Causes of Hypomagnesemia
Common Causes
Grass tetany
Winter tetany
Grass staggers
Calves on a milk-only, magnesium-deficient diet Endurance exercise
Fatty liver syndrome
Uncommon Cause
Undernutrition
■ BOX 22.12
Causes of Hypermagnesemiaa
Epsom salt (MgSO4) overdose given orally or as an enema Intravenous administration of magnesium in excessive amounts Use of MgSO4 IV
aHypermagnesemia is an uncommon condition in large animals.
Disturbances of magnesium metabolism occur principally in cattle and sheep. Complex nutritional and environmental interactions contribute to a variety of clinical syndromes attributed to magnesium deficiency and the onset of tetany in grazing animals.
HYPOMAGNESEMIA. Hypomagnesemia is reported in cattle with grass tetany, transport tetany, winter tetany, and in sheep with grass staggers (Box 22.11) (see also Chapter 41). A serum magnesium level below 1.8 mg/dL (0.75 mmol/L) is considered low; values toxemia
Hypovolemic shock
Acute diarrhea
Colic with strangulated bowel
Strangulating abomasal torsion
Peritonitis
Uroperitoneum (ruptured bladder)
Exercise above anaerobic threshold (normal response in horses)
Uncommon Causes
Renal failure
Renal tubular acidosis
Urea toxicity
Salicylate toxicity
Methanol toxicity
Paraldehyde toxicity
Ethylene glycol toxicity
■ BOX 22.14
Causes of Metabolic Alkalosis
Common Causes
Sequestration of fluid in the abomasum and forestomach in ruminants (internal vomiting for abomasal reflux)
Gastric reflux in horses with ileus
Massive sweat loss in horses (endurance horses)
Chloride depletion
Potassium depletion
Contraction alkalosis (extracellular fluid volume contraction without bicarbonate loss)
Salivary loss of chloride in horses with esophagostomy
Use of diuretics (especially furosemide)
Functional pyloric stenosis
Uncommon Causes
Excessive bicarbonate supplementation or therapy Mineralocorticoid excess
particularly in association with digestive disturbances in ruminants. An initiating process capable of generating alkalosis is necessary and must be coupled with additional factors to maintain metabolic alkalosis. Generation of metabolic alkalosis is traditionally thought to be caused by excessive hydrogen loss, bicarbonate retention, or contraction alkalosis (Box 22.14). Contraction alkalosis occurs when the ECF volume is reduced because of loss or sequestration of fluids high in sodium and chloride but without proportionate loss of bicarbonate, which really contradicts electroneutrality of watery solutions. This is thought to be traditionally a contributing mechanism for the generation of the metabolic alkalosis reported in heavily sweating endurance horses and in response to the diuretic furosemide in the horse. The most common causes of increased hydrogen ion loss are the gastrointestinal losses caused by salivary secretions in ponies after surgical esophagostomy34; massive gastric reflux associated with anterior enteritis, ileus, or small bowel obstruction in horses; and sequestration of fluid in the abomasum and forestomach associated with a variety of gastrointestinal displacements or functional disturbances (vagal indigestion, functional pyloric stenosis) of ruminants. Continuous salivary losses in horses after surgical esophagostomy result in transient metabolic acidosis followed by progressive metabolic alkalosis. Most of these disorders cause significant dehydration and sodium, chloride, and potassium deficits.
The factors responsible for maintaining metabolic alkalosis involve classically explained impaired renal bicarbonate excretion. These factors are associated with the renal response to decreases in the effective circulating fluid volume, chloride depletion, or potassium depletion. Renal tubular sodium resorption is enhanced in response to hypovolemia. Maintenance of electroneutrality requires that sodium resorption in the proximal tubule be accompanied by a resorbable anion, whereas in the distal tubule sodium resorption is associated with the secretion of another cation, usually hydrogen or, to a lesser extent, potassium. Chloride is the only resorbable anion normally present in appreciable quantities in the proximal tubular fluid. In metabolic alkalosis, plasma bicarbonate is increased and the chloride concentration is generally decreased as a result of disproportionate chloride losses. The relative lack of the resorbable anion chloride in the proximal tubule thus allows a larger amount of sodium to reach the distal tubule, where aldosterone and other factors enhance hydrogen or potassium loss into the tubular lumen in exchange for sodium. Potassium depletion reduces or eliminates potassium exchange as a means of sodium retention, thus classically explained placing greater emphasis on hydrogen ion exchange, which stoichio- metrically and quantitatively is difficult to explain. Because renal hydrogen excretion is linked with bicarbonate resorption, the excess bicarbonate cannot be eliminated and metabolic alkalosis is maintained. Thus the traditional explanation is the reason for the paradoxic aciduria seen in some patients with metabolic alkalosis, and it is the reason these patients respond when given intravenous fluids containing sodium, chloride, and potassium. The compensating respiratory response to metabolic alkalosis is hypoventilation, resulting in an increase in the PCO2. Excessive sodium bicarbonate administration is an additional potential cause of metabolic alkalosis. Most normal animals can tolerate large doses of sodium bicarbonate, and excesses are rapidly eliminated by renal excretion.35 However, patients with decreases in effective circulating fluid volume, particularly when coupled with potassium or chloride deficits, may not tolerate a bicarbonate load because renal clearance of excess bicarbonate is likely to be impaired. Attempts to alter the acid-base balance, and thereby affect the athletic performance of racing horses, by prerace administration of high doses of sodium bicarbonate-containing “milkshakes” have become a major concern around the world. This has stimulated substantial research on the acid-base balance of horses before and after racing. In many racing jurisdictions, stringent prerace or postrace standards for blood pH or bicarbonate or TCO2 levels (or both) have been enacted to control this practice. The prerace administration of the diuretic furosemide is allowed in several racing jurisdictions of North America. This drug results in a mild metabolic alkalosis with increased bicarbonate, and allowances for this may be included in some regulations. Racing supplements (which may contain sodium bicarbonate), bicarbonate precursors, and diets that supply a high positive cation-anion balance also have the potential to produce a significant metabolic alkalosis.
Respiratory Acidosis
Respiratory acidosis is characterized by a decrease in pH and an increase in PCO2, which develop because of decreased effective alveolar ventilation. CO2 diffuses through the lungs much more readily than oxygen; thus diseases that compromise ventilation normally result in decreases in PO2 before significant increases in PCO2 develop. The most common causes are acute upper respiratory obstruction and primary pulmonary diseases including pneumonia, pneumothorax, pulmonary edema, and severe equine asthma (Box 22.15). Diseases or drugs that affect
■ BOX 22.15
Causes of Respiratory Acidosis
Common Causes
Primary pulmonary disease Obstruction of the upper airway
Laryngeal edema
Aspiration pneumonia
Pneumonia
Pneumonia and pleuritis complex Pneumothorax
Severe equine asthma
Depression of the respiratory center of the central nervous system
General anesthesia with inappropriately assisted ventilation Drugs
Opiates
Anesthetics
Tranquilizers
Central nervous system diseases
Uncommon Causes
Cardiac arrest
Muscle weakness or dysfunction Tetanus
Botulism
Myasthenia Severe hypokalemia
Neonatal respiratory distress syndrome
■ BOX 22.16
Causes of Respiratory Alkalosis
Common Causes
Hypoxemia
Pulmonary diseases
Congestive heart failure
Severe anemia
Stimulation of the respiratory center of the central nervous system
Psychogenic hyperventilation
Excitement, fear, transport, pain
Gram-negative septicemia
Neurologic disorders
Uncommon Causes
After correction of metabolic acidosis
Inappropriate mechanical ventilation
Salicylate toxicity
the respiratory center of the central nervous system may also produce respiratory acidosis, as can general anesthesia. The compensating response for respiratory acidosis is classically thought to be renal retention of bicarbonate. This response requires days to develop and is seen only in chronic respiratory acidosis. Exogenous sodium bicarbonate does not correct respiratory acidosis and should not be administered to affected patients. It should be treated by improving ventilation and alveolar gas exchange.
Respiratory Alkalosis
Respiratory alkalosis is caused by hyperventilation, which may be stimulated by hypoxemia associated with pulmonary disease, congestive heart failure, severe anemia, or neurologic disorders (Box 22.16) or residence or extended stay at high altitudes. The initial compensating response to acute respiratory alkalosis is a modest decline in the ECF bicarbonate concentration—the result of cellular buffering. Subsequent renal responses result in a decrease in the ECF bicarbonate concentration through reduced renal bicarbonate resorption. The decline in bicarbonate may be offset by chloride retention; thus hyperchloremia and decreased PCO2 may be associated with compensated respiratory alkalosis, as well as with compensated metabolic acidosis. Compensating renal responses for chronic respiratory alkalosis that lasts several weeks actually may be sufficient to return pH to normal.36
Mixed Acid-Base Imbalances
Mixed acid-base disorders occur when several primary acid-base imbalances coexist. Metabolic acidosis and alkalosis can coexist, and either or sometimes both may occur with either respiratory acidosis or respiratory alkalosis. The following factors should be considered when evaluating possible mixed acid-base disorders:
• Compensating responses to primary acid-base disturbances do not result in overcompensation.
• Compensating responses, except for long-term renal compensation, rarely correct pH to normal. A normal pH in a patient with an acid-base imbalance is an indicator of a mixed acid-base disturbance.
• A change in pH in the opposite direction to that predicted for a known primary disorder indicates a mixed disturbance.
• Bicarbonate and PCO2 always deviate in the same direction with primary acid-base disturbances. If these parameters deviate in opposite directions, a mixed abnormality exists.
• If the change in the anion gap does not approximate the change in bicarbonate, a mixed acid-base imbalance should be suspected.
Mixed acid-base abnormalities occur with some frequency in domestic animals and often are overlooked. The practitioner must be aware of the potential for mixed acid-base imbalances to correctly interpret blood gas data in complex clinical situations.
Anion Gap
The anion gap can be an extremely helpful tool for categorizing causal factors in acid-base imbalances and may prove a useful prognostic guide in animals with serious digestive disorders. The anion gap can be calculated as the difference between the major cations (sodium plus potassium) and measured anions (chloride plus bicarbonate). The anion gap normally is 12 to 17 mmol/L and provides an approximation of the so-called unmeasured anions. These are anions that are not measured routinely in the clinical laboratory; they include “physically” the anionic equivalents of plasma proteins (debatably albumin in particular) which comprises about 95% of total anion gap in normal adult animals, and sulfate, phosphate, lactate, and ketones, with a variety of inorganic anions. Significant differences exist in the normal range of the anion gap among species, and there may also be age-related differences. Foals are reported to have a larger anion gap than do adult horses.
Hypoproteinemia and hyperchloremic metabolic acidosis are the most common causes of a decrease in the anion gap. The cause of normal to low anion gap hyperchloremic metabolic acidosis often can be differentiated on the basis of the serum potassium concentration. Animals with hyperchloremic metabolic acidosis associated with gastrointestinal fluid losses or renal tubular acidosis most often manifest hypokalemia, whereas hyperkalemia generally is seen in patients with decreased mineralocorticoid secretion (Addison disease) or renal failure with renal shutdown. Decreases in the anion gap also result from overhydration caused by decreases in the protein concentration and changes in the relative concentration of plasma sodium and chloride.
Most commonly, high anion gap acidosis is associated with accumulation of a metabolizable acid, most commonly L and D lactic acid associated with anaerobic exercise, grain overload, acute gastrointestinal accidents, or hypovolemic shock (e.g., diarrhea in young calves). The commonly used laboratory procedures (mostly automated blood gas machines; point of care equipment) for the determination of lactate only measure the L form of lactate. Microbial fermentation in the gastrointestinal tract may result in the production of both the D and L forms of lactate. Recent studies have shown in milk-fed calves and kids that accumulation of D-lactate is a major factor in the profound acidosis associated with certain digestive disorders.37,38 Unfortunately, at present, determination of D-lactate requires special procedures, which are now available only in special diagnostic commercial kits. Ketoacidosis, uremic acidosis, and poisoning with a variety of anionic poisons result in increases in nonmetabolizable acids that are also causes of an increased anion gap (rare in large animals). When a high anion gap metabolic acidosis is found, a thorough search for the potential causes of the accumulated unmeasured anions is indicated. The anion gap is also useful for identifying mixed acid-base imbalances. A mixed metabolic acid-base imbalance should be suspected when the change in the anion gap does not approximate the change in bicarbonate. Increases in the anion gap can also be associated with dehydration and contraction alkalosis caused by changes in the protein concentration and the relative concentration of plasma sodium and chloride.
Bicarbonate and Total Carbon Dioxide
Bicarbonate accounts for approximately 95% of the measured CO2; thus TCO2 or “CO2 content” of serum or plasma provides a measure of metabolic changes in the acid-base balance. Most automated chemistry profiles now provide the TCO2 level. Bicarbonate or TCO2 is decreased in metabolic acidosis and increased in metabolic alkalosis. However, the bicarbonate or TCO2 values provide only a crude indication of acid-base status. When acid-base abnormalities are suspected, appropriate samples should be submitted for blood gas and electrolyte determination run on the same analyzer. It should be pointed out that bicarbonate levels are always calculated on automated measuring systems.
Buffer Base, Standard Bicarbonate, and Base Excess or Base Deficit
The terms buffer base, standard bicarbonate, and base excess (or base deficit) represent derived calculated estimates of the metabolic component of acid-base balance and do not really physically exist. The buffer base indicates the sum of all the buffer anions in blood under standardized conditions. The standard bicarbonate is the plasma bicarbonate concentration that would be found under specific conditions that eliminate respiratory influences on the values obtained. The base excess or base deficit is often supplied in routine assessment of acidbase balance; it indicates the deviation of bicarbonate from normal, usually the stored “normal” for humans in blood gas machines, and thus not directly applicable to large animal blood gas interpretations. The calculated base deficit provides a means of estimating the amount of bicarbonate required to correct metabolic acidosis. The conundrum is that bicarbonate cannot be given without a cation (mostly Na). The bicarbonate estimate is calculated by multiplying the base deficit by the probable sodium bicarbonate space (about 30% [adults] to 50% [neonates and young animals] of body weight), as in the following equation:
mEq NaHCO3- needed
= mEq base deficit ? kg body weight ? 0.3 or 0.5
It should be pointed out that normal ranges of plasma bicarbonate concentrations are different in humans and large animals, with normal large animals being an average of 4 to 5 mEq/L higher compared with normal humans. Modern blood gas machines providing base excess calculations most commonly will use human equations with normal bicarbonate concentrations of 24 mEq/L. When given clinical interpretations of values of base excess, this should be taken into consideration when applying the above equation to calculate NaHCO3- deficits. The factor of 0.3 to 0.5 actually represents the sodium space in adult and young animals encompassing the ECF compartment.
STRONG ION-CENTERED APPROACH TO ACID-BASE BALANCE.
In the strong ion-centered approach (also known as the Stewart approach or physicochemical approach), acid-base balance is determined by three independent variables: strong ion difference (SID), the partial pressure of CO2, and the total concentration (mass) of nonvolatile weak acids (Atot), the principal components of which are plasma proteins and, to a lesser extent, inorganic phosphate.33,39 Bicarbonate and pH are dependent variables determined by these three independent variables. Briefly, this approach is a quantitative approach applying physical laws of chemistry of watery solutions to calculate parameters involved in determining acid-base equilibria and disequilibria. These laws include electroneutrality (Σ cations = Σ of anions), dissociation equilibria (proteins, bicarbonate system, water), and mass conservation. If one of the independent variables (strong electrolytes, proteins, or PCO2) changes the other dependent variables, pH, bicarbonate, and weak electrolytes adjust. Several studies have adapted the physicochemical approach for practical application in human and veterinary medicine.32,39 44 Experimentally determined species-specific Atot and Ka data are now available for horses, cattle, dogs, and humans.45-48
Strong Ion Difference
Strong electrolytes are completely dissociated in aqueous solution and chemically nonreactive. The SID is simply the difference between the total concentration of strong cations (sodium, potassium, calcium, and magnesium) and total concentration of strong anions (chloride, sulfate, lactate, acetoacetate, and β-hydroxybutyrate) with sodium, potassium, and chloride normally being quantitatively the principal determinants of SID. In fluids with little or no protein, such as cerebrospinal fluid (CSF), bicarbonate concentration is the same as the SID, but it should be noted that the chloride concentration is higher (≈10 to 11 mEq/L) and SID is lower compared with blood; thus normal acid-base equilibria in such fluids are different from acid-base equilibria of blood. Abnormalities in PCO2 are viewed in essentially the same manner in both the traditional and nontraditional approaches to acid-base balance. Plasma albumin debatably makes up the majority of Atot. The Atot (total mass of proteins) exists physically in blood in both dissociated (A-) and undissociated forms (HA). A decrease in Atot (A-) resulting from hypoalbuminemia/hypoproteinemia causes an alkalosis with an increase in bicarbonate, whereas hyperalbuminemia has the opposite effect. Increases in A- can cause an increase in anion gap, whereas decreases in A- result in a decrease in anion gap. A change in protein concentration has effects on acid-base balance. The literature describes hyperproteinemic acidosis or hypoproteinemic alkalosis.40 Details are outlined in Figs. 22.2 and 22.3.
When protein and inorganic phosphate remain within the normal range, acid-base balance is controlled by changes in PCO2 mediated by the respiratory system and changes in SID, which are largely under the control of the kidneys. Heavy sweat loss in an endurance horse, displaced abomasum in a cow (trapping of chloride in abomasum), or the prerace administration of the diuretic furosemide (chloruresis) in a racehorse produces metabolic alkalosis (mainly observed abnormalities are decreases in serum chloride concentrations). In each case the alkalosis is the result of a disproportionate loss of chloride relative to sodium, yielding hypochloremia and an increase in SID. The
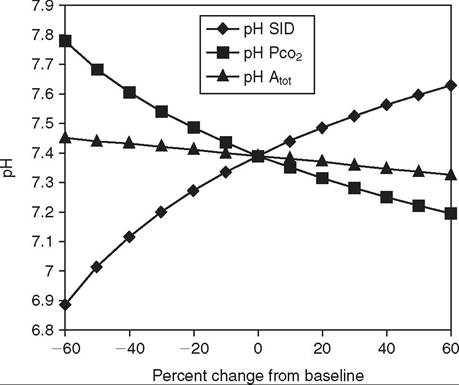
FIG. 22.3 Spider plot graphically showing quantitatively the dependence of plasma pH concentration on changes in the three independent variables (SID, PCO2, and Atθt). The spider plot was generated by systematically varying one independent variable while holding the other two independent variables at their reference values (y axis values at 0 on x axis) for plasma.
dissociation equilibrium of serum bicarbonate concentrations will adjust to fulfill electroneutrality requirements of watery biologic solutions. Correction of the alkalosis is brought about by the provision of relatively more chloride relative to sodium, generally as sodium chloride or potassium chloride. This results in changing SID to more normal concentrations and thus a return of bicarbonate and pH toward normal. Metabolic acidosis with a large base deficit is generally treated with sodium bicarbonate, with sodium being the strong cation added to the ECF and bicarbonate adjusting according to its dissociation equilibrium and sodium added without a strong anion. Other metabolizable anions (short-chain fatty acids such as lactate, acetate, gluconate, and propionate, added with Na as the cation) could be substituted for bicarbonate and achieve similar effects. Short-chain fatty acids are quickly metabolized to glucose and ATP as sources of energy for the body; in physicochemistry, this will result in a relative increase of the SID.
Calculation of SID is simple and provides useful insight in patients with metabolic acid-base disturbances. Factors that influence SID range from changes in free water, to sodiumchloride imbalances that result from excessive losses or disproportionate retention of sodium or chloride, to the accumulation of strong organic anions (most commonly L and D lactate). Organic acidosis can be produced by the accumulation of exogenous and endogenous anions. The anion gap does not always accurately predict the presence of unmeasured strong anions. Mathematic methods have been developed as a means for the detection of unmeasured anions49,50 and for the calculation of the simplified strong ion gap.51
Both the traditional and nontraditional or strong ion approaches to acid-base balance have proven useful to address practical problems in both research and medical settings. The traditional approach to acid-base balance is more widely accepted and user-friendly. The strong ion approach may provide a better understanding as to why the bicarbonate concentration is changing as it integrates acid-base and electrolyte disorders. The strong ion approach has recently gained wider acceptance from members of the human critical care community, who have found it useful for the analysis of the complex fluid, electrolyte, and acid-base problems of patients in intensive care units. Several easy-to-use computer or calculator programs have been developed for the mathematically challenged that facilitate implementation of the strong ion approach to acid-base balance.
Serum Enzymes
Serum enzyme levels are proportional to their cell or tissue concentrations and activity. A number of variables affect the appearance in serum or plasma. The amount of any single enzyme in serum depends on concentration in the cell, total tissue mass, magnitude of cell/organ damage, normal death of the cell, apoptosis, degradation rate of the enzyme in serum or its half-life, and whether the injury is single or continuous.2 Some of the common and less common causes of elevated serum enzyme activity are listed in Box 22.17.
Sorbitol Dehydrogenase
Sorbitol dehydrogenase (SDH) is a cytoplasmic enzyme that reversibly oxidizes D-sorbitol to D-fructose with the cofactor NAD. It is a liver-specific enzyme in all large animal species. Increases in this enzyme indicate hepatocellular damage and leakage of enzymes. Increases in SDH are also seen with obstructive or strangulating gastrointestinal lesions and with acute toxic enteritis as a result of liver damage associated with absorption of bacteria or their toxins (or both) from the damaged bowel into the portal circulation. This enzyme is a sensitive indicator of liver damage, and modest increases may be seen with anoxia, acute anemia, or general anesthesia. The half-life of SDH in the circulation is short (a matter of hours), and elevations indicate active and ongoing liver damage. This enzyme is not stable when stored at room temperature (only for 5 hours), but refrigerated samples may yield useful results up to 24 to 72 hours if frozen. SDH activity has also been observed increased in horses and cattle with intestinal diseases and in foals with sepsis indicating hepatic involvement in these circumstances.52,53
Creatine Kinase
Creatine kinase (CK) is present in high concentrations in the cytoplasm of myocytes; it catalyzes the reversible phosphorylation of creatinine to form creatinine phosphate, which is a major storage of high-energy phosphate in the muscle. It is a “leakage” enzyme and a highly sensitive and specific indicator of muscle damage in domestic animals. Normal values of CK activity vary according to physical activity, restraint, age, and sex. There are four important CK isoenzymes. CK-1 is a BB isoenzyme found mostly in brain. CK-2 MB isoenzyme is found mainly in cardiac muscle and small proportions in skeletal muscle. CK-3 MM isoenzyme is found mainly in skeletal muscle and lesser levels in heart muscle and is responsible for the serum CK levels in large animals. CK-Mt is found mainly in mitochondria and is 15% of cardiac activity.54 Although CK is found in both cardiac and skeletal muscle, elevations of this enzyme most commonly are associated with exertional myopathies (rhabdomyolysis) and are seen as musculoskeletal manifestations of systemic disease. Intramuscular injections, vigorous exercise, or prolonged shipping may result in modest releases (up to a fourfold increase over resting values) of CK into the circulation without producing histologic evidence of muscle damage. Endurance exercise may lead to moderate elevation of CK (2000 to 15,000 IU/L) in some horses that show no recognizable sign of exertional myopathy. Experimentally it has been shown in horses that when administered IV, the half-life of this enzyme in the circulation is short, 2 hours, but if given IM the half-life is 12 hours.55 In cattle the half-life is 8 to 9 hours56; its activity peaks at 6 to 12 hours, and even marked elevations in CK may return to normal within 24 hours after a single muscle insult. Although marked elevation of CK can be a guide to the extent of muscle damage, the short half-life and the potential for continuing myonecrosis have a marked influence on the enzyme activity observed at any point in time. A persistent elevation of CK suggests a process resulting in active and continuing muscle damage and provides grounds for
■ BOX 22.17
Causes of Elevated Serum Enzymes
Elevation of Sorbitol Dehydrogenase (SDH)
Common Causes
Severe anoxia
Acute liver failure
Liver abscess
Secondary to damaged bowel Strangulating intestinal lesion Acute toxic enteritis
Chronic liver failure
Less Common Causes
Acute and severe anemia
General anesthesia
Anoxia
Elevation of γ-Glutamyltransferase (GGT)
Common Causes
Acute liver failure
Chronic liver failure
Pyrrolizidine alkaloid toxicity
Aflatoxicosis
Cholangiohepatitis
Cholelithiasis
Liver flukes
Uncommon Causes
Higher normal range in young animals Fatty liver
Elevation of Alkaline Phosphatase (ALP)
Common Causes
Acute liver failure
Chronic liver failure
Pyrrolizidine alkaloid toxicity Cholangiohepatitis
Cholelithiasis
Liver flukes
Uncommon Causes
Higher normal range in young animals Fatty liver
Elevation of Creatine Kinase (CK)
Common Causes
Exertional rhabdomyolysis (azoturia, myositis, tying-up) Polysaccharide storage myopathy
Streptococcus equi-associated myopathy
Nutritional myodegeneration (selenium, vitamin E deficiency) Post-endurance ride multisystemic disorder
Alert downer cow syndrome (muscle crush syndrome) Malignant hyperthermia
Malignant edema
Prolonged recumbency with inability to rise
Uncommon Causes
Normal postexercise modest increase Acute cardiomyopathy
Purpura hemorrhagica
Equine influenza
Sarcosporidiosis
Local irritation from intramuscular injections
Elevation of Lactate Dehydrogenase (LDH)
Common Causes
Muscle disease
Exertional rhabdomyolysis (azoturia, myositis, tying-up)
Polysaccharide storage myopathy
Streptococcus Equi-Associated Myopathy
Nutritional myodegeneration (selenium, vitamin E deficiency) Post-endurance ride multisystemic disorder
Alert downer cow syndrome (muscle crush syndrome) Malignant hyperthermia
Malignant edema
Liver disease
Acute liver failure
Chronic liver failure
Cholangiohepatitis Cholelithiasis
In vitro hemolysis
Uncommon Causes
Hemolytic anemia
Acute cardiomyopathy
Purpura hemorrhagica
Equine influenza Sarcosporidiosis
Local irritation from intramuscular injections
Fatty liver
Elevation of Aspartate Aminotransferase (AST)
Common Causes
Muscle disease
Exertional rhabdomyolysis (azoturia, myositis, tying-up)
Polysaccharide storage myopathy
Streptococcus equi-associated myopathy
Nutritional myodegeneration (selenium, vitamin E deficiency) Post-endurance ride multisystemic disorder
Alert downer cow syndrome (muscle crush syndrome) Malignant hyperthermia
Malignant edema
Liver disease
Acute liver failure
Chronic liver failure
Cholangiohepatitis
Cholelithiasis
Liver flukes
In vitro hemolysis
Uncommon Causes
Hemolytic anemia
Acute cardiomyopathy
Purpura hemorrhagica
Equine influenza Sarcosporidiosis
Local irritation from intramuscular injections
Fatty liver
resting athletic horses; it can also be used as a prognosticator in downer cows (>3500 IU/L carries a guarded prognosis). Elevated CK provides no information on the factors responsible for the rhabdomyolysis; more precise information regarding the etiology can be obtained by muscle biopsy and its histologic and histochemical examination. Hemolysis or delayed separation of serum from clot may produce falsely high values for CK as a result of inadvertent muscle penetration during venipuncture.
Aspartate Aminotransferase
Aspartate aminotransferase (AST) is found in high concentrations in a variety of tissues, including skeletal and cardiac muscles, the erythrocytes and kidneys, and the liver. AST enzyme catalyzes the transfer of the α-amino group of aspartic acid to α-ketoglutaric acid, resulting in formation of oxaloacetic acid and glutamic acid. This enzyme is a nonspecific indicator of tissue damage and tends to be less sensitive to mild insults than the tissue-specific enzymes SDH and CK. The half-life of AST in the circulation is relatively long (it has been reported in horses to be between 7 and 8 days by some and 3 to 4 days by others57 and around 1 day in cattle58), and elevations may persist for as long as 10 days after an episode of myonecrosis or liver damage. As a general rule, extensive muscle necrosis tends to produce much higher elevations of AST than severe liver necrosis. This enzyme is most useful when compared with the tissue-specific enzymes as determined sequentially over the time course of a disease process. Elevations of CK and AST indicate muscle damage, whereas elevations of SDH and AST indicate liver damage. Marked but transient elevations of CK and SDH are associated with a single insult to the muscles and liver, respectively, whereas AST increases gradually and remains elevated for a much longer time. Thus a moderate to marked increase in AST in an animal with progressively declining SDH or CK indicates that some tissue damage occurred within the past 7 to 10 days, but also that the process may no longer be active. This is often a favorable prognostic indicator. Persistent elevation of or a progressive increase in CK or SDH and AST over time indicates an active, continuing process of tissue damage, and the prognosis is more guarded. AST is relatively stable at room temperature, but hemolysis, icterus, or lipemia may interfere with the assay.
Y-Glutamyltransferase
Y-Glutamyltransferase (GGT) is a membrane-bound enzyme that functions in the γ-glutamyl cycle, where it catalyzes the transfer of γ-glutamyl groups from γ-glutamyl peptides. It is important in glutathione metabolism, amino acid absorption, and protection against oxidant injury. This enzyme is found in the biliary tract from where it derives the serum activity in health and disease, therefore making it an important marker of hepatobiliary disorders and cholestasis in large animals. GGT is also found in the pancreas, gastrointestinal tract, urinary tract, mammary gland, and reproductive tract; it does not increase in serum in disorders involving these organs. GGT is quite stable, and reliable results can be obtained from samples submitted several days after blood samples have been drawn, provided the serum is refrigerated. The half-life of GGT activity in blood is not known, but it has been suggested to have a half-life of 3 days in horses. The activity of this enzyme is highest in the cells of the periportal region of the liver, in the pancreas, and in the renal tubular cells. Pancreatic diseases resulting in inflammation and necrosis are relatively rare in large animal species. Damage to the renal tubular cells leads to a release of GGT into the tubular lumen and urine. Because this enzyme is a relatively large molecule, it is not resorbed into the systemic circulation and renal tubular damage does not result in elevated serum GGT activity. Increases in GGT relative to creatinine in the urine have been used as an index of acute renal tubular damage. However, the validity of the normal range for this ratio in horses has been questioned.
In large animal species, an elevation in serum GGT is one of the more reliable indicators of damage to the liver and biliary obstruction. Disease processes such as pyrrolizidine alkaloid intoxication (in subclinical liver diseases showed a 75% sensitivity and 90% specificity); chronic active hepatitis, cholangiohepatitis, and cholelithiasis produce liver damage, primarily in the periportal region, leading to marked and persistent elevation of GGT activity in the serum.59 In these instances, elevations in serum ALP activity are generally associated with the increase in GGT. Two syndromes—fatty liver syndrome in dairy cows and hyperlipemia syndrome of periparturient mares of pony and miniature horse breeds—are associated with liver damage, which is often reflected by elevation of GGT.
Most suckling neonatal large animals, except horses, have high levels of GGT activity in their serum. This is the result of absorption of maternal GGT, which is present in relatively high levels in the colostrum, making its serum levels a sensitive indicator of passive transfer. Elevation of GGT in neonates should be regarded as a normal finding unless it is associated with other evidence of liver disease. Serum GGT concentrations are relatively higher in foals than in adults.60 The normal range of serum GGT activity for burros, donkeys, and asses may be substantially higher (two to three times) than the normal range of serum GGT activity for horses. Caution should therefore be used when evaluating this enzyme in these species. Elevation in serum GGT activity has been reported in Thoroughbred racehorses that are performing below expectations. The reasons for the increase in GGT are not known, but horses often respond to a period of rest or reduction in workload. These horses show little histologic evidence of liver damage, and other indices of liver damage and dysfunction usually are within normal limits. The stress of training may be associated with an elevated GGT. Certain trainers, often highly successful ones, appear to have a disproportionately large number of horses with elevations of this enzyme. The normal range for GGT of Thoroughbreds in race training may be slightly higher than that of normal sedentary horses.
Alkaline Phosphatase
Alkaline phosphatase (ALP) is used in most species as a marker for intrahepatic or extrahepatic obstruction of the biliary system. The enzyme is also released by osteoblasts from metaboli- cally active bone. This may be the reason that young, rapidly growing animals normally have high levels of serum ALP, which accounts for the 100-fold higher levels in foals than in adults. Serum ALP in adult horses is 80% liver ALP and 20% bone ALP. Elevations in ALP are also reported in cases of rickets and healing fractures. The intestinal isoenzyme of ALP is similar to the ALP isoenzyme found in neutrophils. Elevations in the ALP activity of abdominal fluid in horses with intraabdominal disease may reflect the release of this enzyme from the neutrophils rather than being a specific marker of damage to the bowel.
ALP has been useful for evaluating liver disease in large animals, particularly in horses with pyrrolizidine alkaloid intoxication, chronic active hepatitis, and cholangiohepatitis, and in some patients with cholelithiasis. A profound elevation in ALP activity is thought to reflect periportal liver damage and biliary obstruction in these patients. A moderate to marked elevation in ALP may be observed in a wide range of disorders resulting in hepatic necrosis and intrahepatic cholestasis. Because this enzyme is not organ specific in large animals, elevations in ALP activity must be interpreted in relation to more organspecific enzymes such as SDH and GGT.
Other Enzymes
LDH is a metalloenzyme that hydrolyzes many types of phosphate esters at an alkaline pH in the presence of zinc and magnesium ions. It is found in relatively high concentrations in a variety of organs and tissues of the body from the heart, liver, muscle, and kidney to the erythrocytes and leukocytes. An elevation in serum LDH enzyme activity must be evaluated in relation to other, more organ-specific enzymes. LDH isoenzyme analysis can be helpful in differentiating organ system damage, but the analysis is time consuming and not always available. An elevation in LDH activity is expected in hepatic necrosis and serves as an indicator of an active disease process. Extensive muscle damage and rhabdomyolysis tend to result in a more massive release of enzyme and much higher serum enzyme activity. A modest elevation in LDH may be seen in some hemolytic disorders and some cases of leukemia. Blood samples must be handled with care because hemolysis results in falsely elevated serum LDH activity.
Glutamic dehydrogenase (GLDH) and ornithine carbamoyltransferase (OTC) are two enzymes that are reported to be sensitive indicators of hepatic necrosis in ruminants. GLDH is a mitochondrial enzyme that catalyzes the conversion of glutamate to 2-oxoglutarate. Injury to the hepatocyte has to be severe enough to damage the mitochondria. It is suggestive of either hepatocyte death or sublethal hepatocyte injury. In large animals, it is more liver specific than AST and has better storage stability than SDH. OTC is a hepatic- sensitive enzyme that catalyzes the reaction between ornithine and carbamoylphosphate-forming citrulline and inorganic phosphate. OTC has been used mostly to evaluate fatty liver in cattle and as a prognostic indicator in abomasal displacement and fatty liver.61 ALT is an important liver-specific enzyme that has wide application in small animals and is often included in automated chemistry profiles. This enzyme has not been useful for evaluation of liver disease in large animal species, and occasionally horses with marked rhabdomyolysis and no other evidence of liver disease show an elevation in serum ALT activity.