Phylogenetic Position of Zoophilic Dermatophytes and Their Identification
The dermatophytes have been divided into three genera by using conventional phenotypic taxonomy: Trichophyton, Microsporum, and Epidermophyton. This classification scheme has been widely accepted in the second half of the twentieth century and used till today.
Phylogenetic relationships between dermatophytes have been exploited by using numerous genetic loci, including mitochondrial DNA (mtDNA) (Nishio et al. 1992), small subunit of ribosomal DNA (SSU rDNA) (Harmsen et al. 1995), large subunits (LSU) of rDNA (Leclerc et al. 1994), internal transcribed spacer (ITS) region of rDNA (Graser et al. 1999a, b, 2000b; Makimura et al. 1999; Summerbell et al. 1999; Kawasaki et al. 2011; Pchelin et al. 2016), chitin synthase (CHS) (Kano et al. 1999, 2002; Hirai et al. 2003) and DNA topoisomerase II genes (TOP II), actin, glyceraldehyde-3-phosphate dehydrogenase (GPD) (Kawasaki et al. 2011), translation elongation factor 1-α (TEF 1-α) (Mirhendi et al. 2015), β-tubulin (Rezaei-Matehkolaei et al. 2014; Pchelin et al. 2016), and calmodulin (Ahmadi et al. 2016). The topology of phylogenetic trees based on ITS region (Fig. 3.3) is congruent with those based on protein-coding loci, i.e., TEF 1-α, β-tubulin, and calmodulin. The studies with sufficient coverage of species across diversity of Arthrodermataceae consistently showed that both Trichophyton and Microsporum are polyphyletic prompting reevaluation of the generic concept of dermatophytes (Graser et al. 1999a; Rezaei-Matehkolaei et al. 2014; Mirhendi et al. 2015; Ahmadi et al. 2016). The new generic concept with monophyly as a main criterion was recently introduced by de Hoog et al. (2017) who increased the number of recognized genera from three to seven. This new concept has only limited consequences for the taxonomy of human and animal pathogenic dermatophytes, while significant changes were made in the taxonomy of geophilic species.ITS rDNA sequencing is considered a gold standard for species identification. It is still the only genetic locus available for all currently accepted species, is recommended as a barcode, and is widely used for identification in praxis (Irinyi et al. 2015). ITS region sequence has capability to discriminate even closely related species, e.g., M. canis, M. audouinii, and M. ferrugineum; T. benhamiae and T. concentricum; and T. quinckeanum and T. schoenleinii. Identification of all mentioned sibling species cannot be realized by amplification of any currently available protein-coding loci. When DNA sequence is available, species identification is usually obtained by comparison with the reference sequence in the database (e.g., GenBank; www.ncbi.nlm.nih.gov/genbank). Because GenBank acts primarily as an archive, many sequences have been annotated incorrectly, the reason why curated databases were established, e.g., ISHAM ITS Database (http://its. mycologylab.org) or Dermatophytes Species Database at Westerdijk Fungal Biodiversity Institute (http://www.westerdijkinstitute.nl/Dermatophytes). A plenty of PCR-based techniques were developed for identification of dermatophytes with variable discriminatory power including nested PCR, PCR-RFLP, RAPD, ISSR- PCR, AFLP, AP-PCR, etc. (Chen et al. 2011; Sharma and Graser 2011; Cafarchia et al. 2013c). In particular PCR-RFLP targeting ITS or other regions with digestion using various restriction enzymes reached relatively broad popularity due to low financial cost and usability for identification of major pathogens (De Baere et al. 2010; Heidemann et al. 2010; Rezaei-Matehkolaei et al. 2013). Many other methods showed similar or higher potential for species identification, but they have only limited usability due to poor reproducibility or laboriousness and are now obsolete.
While DNA-based molecular methods are usually highly accurate and rapid, they are relatively costly and can be complex to implement. Matrix-assisted laser
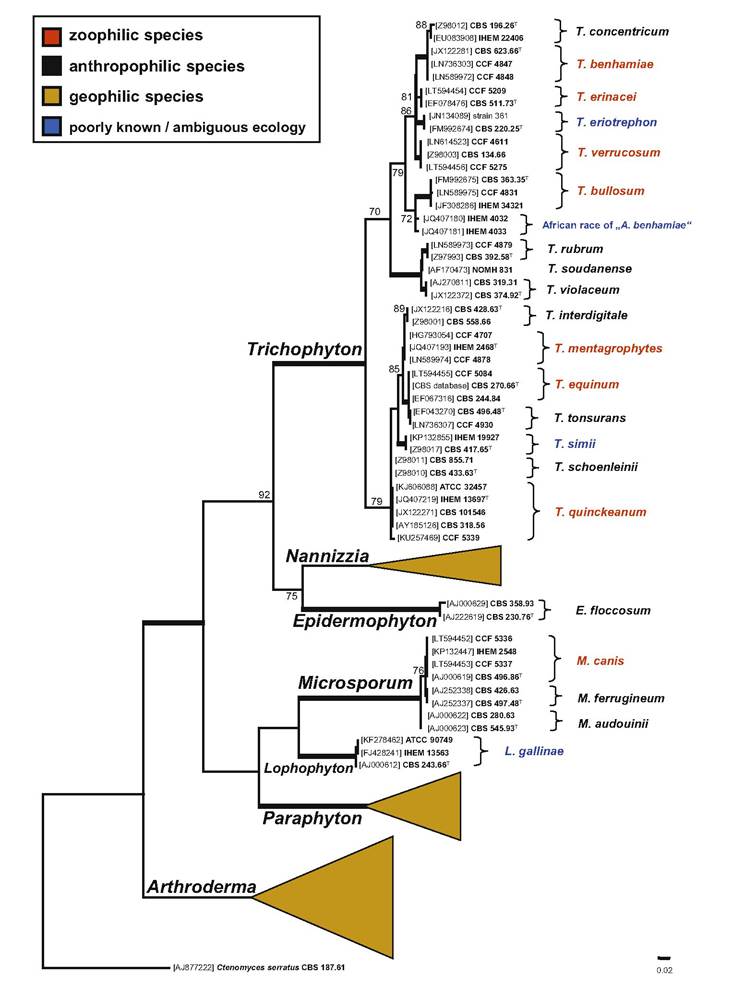
Fig.
3.3 Phylogenetic placement of zoophilic species within dermatophytes. Best scoring maximum likelihood tree-based ITS rDNA constructed with the IQ-TREE version 1.4.0 (Nguyen et al. 2015) by using GTR + I + G4 substitution model. Dataset contained 95 taxa and a total of 731 characters of which 351 were variable and 306 parsimony informative. Support values atdesorption ionization time-of-flight mass spectrometry (MALDI-TOF MS) may represent an alternative to conventional dermatophyte identification. Application of MALDI-TOF MS as an identification procedure for pathogenic dermatophytes has been increasingly used in the laboratories due to its time- and cost-effectiveness (despite the initial cost of investment for obtaining the equipment) (L'Ollivier and Ranque 2017). The method is able to distinguish all major pathogenic dermatophytes; however, the differentiation of phylogenetically closely related taxa is associated with higher level of errors, e.g., T. quinckeanum and T. schoenleinii, different races of T. benhamiae, or T. interdigitale and T. mentagrophytes. Thus, supplementation of the reference spectra libraries is still required for optimal dermatophyte identification (Graser 2014).
Despite the advent of molecular methods, conventional species identification of dermatophytes is still the prevailing method of identification worldwide and consists of micro- and macromorphological examination of cultures, sometimes supplemented with various physiological and biochemical tests (e.g., nutritional tests, vitamin requirements tested on T1-T7 Trichophyton agars, urease activity, hair perforation test, etc.), mating experiments, etc. The Sabouraud's agar, frequently supplemented by antibiotics chloramphenicol and cycloheximide, is the most commonly used isolation medium. Strains with typical morphology can be identified directly from primary cultures, but subculturing on specific media inducing sporulation or production pigments may be necessary (Robert and Pihet 2008).
3.6