Specific Diseases of the Nervous System
Viral and Prion Diseases
Rabies
Though the occurrence of rabies in goats is infrequent, rabies should not be left out of the differential diagnosis of caprine neurologic disease for two reasons: rabies is a consistently fatal zoonotic disease; and the signs of rabies are highly variable and may mimic other neurologic conditions.
Etiology
Rabies virus is a bullet-shaped, enveloped, single-stranded, negative-sense, non-segmented RNA virus in the genus Lyssavirus of the family Rhabdoviridae of the order Mononegavirales. The viral genome encodes for five proteins: nucleoprotein, phosphoprotein, matrix protein, glycoprotein, and an RNA polymerase. The lyssaviruses are neurotropic viruses, well adapted to replication in the mammalian nervous system. There are currently 17 species recognized in the genus Lyssavirus, divided among at least three phylogroups (ICTV 2017).
With the exception of rabies virus, which occurs essentially worldwide, these lyssaviruses have distinct but sometimes overlapping geographic distributions. Australian bat lyssavirus (ABLV), for example, is found solely in Australia. The European bat viruses are distributed among insectivorous bats in Eurasia. Lagos bat virus, Duvenhage virus, and Mokola virus have not been encountered outside of Africa (Markotter et al. 2006). Neutralizing antibody to Mokola virus has been identified in goats in Nigeria (Kemp et al. 1972; Nottidge et al. 2007).
Within the rabies virus genotype, virus variants associated with particular host species and geographic areas are recognized. They can be identified by reactions with panels of monoclonal antibodies or by genetic analysis of nucleotide substitutions (Rupprecht et al. 2002). In the continental United States, for example, there are three distinct rabies virus variants infecting skunks, found in California, the upper Midwest, and the lower Midwest, while a raccoon-adapted variant predominates on the eastern coast.
Two gray fox-adapted variants are found focally in Arizona and Texas and separate variants adapted to red foxes and arctic foxes are found in Alaska. In addition, however, there are numerous independent reservoirs of rabies variants in insectivorous bats, whose overlapping ranges essentially make the entire United States enzootic for rabies (Blanton et al. 2006).The rabies virus grows in tissue culture, embryonating chicken eggs, and suckling mice. It has a short survival time outside the mammalian host and it becomes non-viable in dried saliva within a few hours. It is a relatively fragile virus, readily inactivated by ultraviolet irradiation and common disinfectants. For this reason, thorough cleansing and disinfection of bite wounds immediately following their occurrence constitute a very important and useful aspect of rabies prophylaxis.
Epidemiology
Several countries, islands, and regions are rabies free, including Antarctica, Australia, New Zealand, Scandinavia, United Kingdom, Ireland, Cyprus, Japan, Papua New Guinea, and Hawaii, but remain vigilant against the introduction of infected animals. However, in much of the world rabies is still enzootic. In Europe and North America most reported rabies is sylvatic, occurring primarily in wildlife reservoirs such as foxes, skunks, raccoons, and insectivorous bats. In much of Asia, Africa, and the Middle East rabies is still largely urban, with dogs and cats comprising the majority of cases. In Latin America rabies is an important problem, especially in dogs and cattle, with bites by vampire bats the primary source of livestock infection. Though Australia is still considered to be free of rabies, the related lyssavirus, Australian bat virus, was identified in the mid-1990s and has produced fatal encephalitis in several people. Though the virus has been identified in numerous species of Australian bats, there is no evidence of infection or spillover of ABLV into terrestrial mammal populations. Nevertheless, animals bitten or scratched by infected bats may contract disease, as evidenced by the confirmation of ABLV infection in two horses with neurologic disease in Queensland in 2013 (WHA 2019).
Rabies remains a serious zoonotic disease worldwide, particularly in developing countries of Asia, the Middle East, and Africa, where comprehensive programs to control rabies in dogs may be lacking and access to or awareness of postexposure prophylaxis is limited. Worldwide, there are an estimated 59 000 human deaths due to rabies each year (WHO 2020a). India alone accounted for up to 60% of the world's rabies cases annually during the 1990s, although improved public education and greater availability of postexposure prophylaxis have improved the situation somewhat in recent years (Sudarshan et al. 2007). In the developing countries, domesticated dogs continue to be the major reservoir of rabies and dog bites the major cause of human cases. Dog bites are most likely the major cause of goat cases as well.
Goats are moderately susceptible to rabies, in the same category with sheep, cattle, and dogs (Crick 1981). While outbreaks in goats are uncommon, goat rabies does occur sporadically in enzootic regions. In the United States in 2018, for example, there were 4951 cases of rabies reported in animals. Wild animals accounted for 4589 (92.7%) of these cases and domestic animals 362 (7.3%). None of these domestic animal cases occurred in goats (Ma et al. 2020). By contrast, in 2005 there were 6 confirmed goat rabies cases in the US out of a total of 5923 confirmed cases in wildlife and 494 in domestic animals (Blanton et al. 2006). Among 34 countries of Europe, including the Russian Federation, Ukraine, and Turkey, there were 4464 rabies cases documented in 2018. Wildlife other than bats accounted for 1597 (35.8%) and domestic animals for 2816 (63.1%) of these cases. Goats and sheep, reported together, accounted for only 106 of the domestic animal cases (WHO 2020b).
In the United States the principal reservoirs of sylvatic rabies are raccoons (Procyon lotor) on the east coast, skunks (mainly Mephitis mephitis) in the Midwest and California, and foxes (mainly Vulpes vulpes) along the Texas-Mexico border.
However, insectivorous bat reservoirs have major public health significance. From 1900 through 2005, 48 human rabies cases were confirmed in the United States, with 10 infections originating outside the country. Of the remaining 38 cases, 35 (92.1%) were infected with bat rabies variants (Blanton et al. 2006).In Europe the principal reservoir is the red fox (V vulpes). The raccoon dog (Nyctereutes procyonoides) has been growing in significance in recent years, especially in the Baltic states of northeastern Europe as the raccoon dog population migrates westward from Asia (Holmala and Kauhala 2006). Oral rabies vaccination campaigns targeting red foxes have been extremely effective, with many countries of western and central Europe declaring rabies- free status in recent years. The continued expansion of the raccoon dog population in Europe therefore is a major concern.
In tropical portions of Mexico and in northeastern Brazil, cases of goat rabies have been associated with vampire bat bites (Batalla et al. 1982; Silva and Silva 1987; Shoji et al. 2006). In Nigeria, where rabies continues to be a serious problem in humans and dogs and goat numbers are high, only eight goat cases were documented nationwide over 33 years. All were presumed to be associated with dog bites, except one attributed to a genet bite (Okoh 1981). In Bombay, India, 3% of 265 animal rabies cases diagnosed in 1981 and 1982 were goats maintained in urban settings and bitten by dogs (Jayarao et al. 1985).
Pathogenesis
All mammals are susceptible to rabies infection. Though rabies can be transmitted rarely by aerosol infection of mucous membranes, rabies virus is most likely introduced into the goat by the bite of a rabid animal whose saliva is infected with the virus. Replication of the virus occurs initially in muscle at the site of introduction and the infection may remain localized at the initial site for many days. The virus then enters local nerves and moves centripetally up nerve trunks to the CNS.
Variations in incubation times and clinical presentation are largely a function of the distance the virus must migrate to the CNS and whether it first reaches the brain or spinal cord. When an animal is bitten on the face, the incubation time tends to be shorter and the clinical presentation is one of encephalitis. When an animal is bitten on the hindlimb, there is a longer incubation period and the early clinical signs suggest an ascending myelitis. Wherever it first reaches the CNS, the virus continues to replicate and extend throughout the brain and spinal cord. At the same time, it begins to move centrifugally back down nerve trunks to nerve endings. In this manner, virus becomes present in saliva via nerve endings in the salivary gland. Hematogenous spread of rabies is extremely rare.When canine-derived virus was injected into the masseter muscles of experimental goats, incubation periods for clinical rabies ranged from 14 to 24 days (Umoh and Blenden 1982). Immunofluorescence studies indicated that virus was abundant in brain tissues, but little or no virus was evident in distal sections of cranial nerves. Whether this low concentration of virus in peripheral nerves was a function of the experimental design or an indication that goats may not shed virus extensively during clinical rabies is unclear. No documented cases of human rabies resulting from exposure to goats could be found in the literature during preparation of this text.
Clinical Findings
Clinical reports of rabies in goats are rare. A 10-year retrospective of rabies in South Africa identified 18 cases of caprine rabies. The furious form appears more commonly than the dumb (paralytic) form in goats. Aggressive behavior was the most common sign, occurring in 83% of cases. Excessive bleating was observed in 72% of cases, salivation in 29%, and paralysis in only 17%. In no case was hydrophobia, straining, pica, or circling observed (Barnard 1979). The clinical course is usually between one and five days and always results in death.
In Brazil, an outbreak of the paralytic form of rabies was documented in goats (Moreira et al. 2018). Six Saanen goats in a herd of 47 died over a period of 15 days. These goats showed signs of apathy, isolation from the herd, and sternal recumbency progressing to lateral recumbency with paddling movements followed by death. The course of illness for each was three to five days. A seventh goat isolated from the herd and was examined by a veterinarian. It was tachypneic and febrile. Over the following 12 hours, the goat developed signs of depression, somnolence, auricular hyperesthesia, opisthotonos, and severe ataxia. After another 8 hours, the goat was in lateral recumbency, paddling, and vocalizing, and was euthanized. A definitive diagnosis of rabies was made at necropsy by direct fluorescent antibody (DFA) test and mouse intracerebral inoculation test (MIT). The owner reported a history of bat bites in his livestock.
In experimental caprine rabies, early signs observed in 19 goats were variable and included hyperexcitability, apparent blindness, anxiety, aggressiveness with piloerec- tion, pruritus with self-mutilation, salivation, frothing, depression, standing off in corners, shaking of the head, and fine tremors, especially in the thighs. Later signs included frothy salivation, which was common, incoordination, staggering, circling, torticollis, extensive muscle tremors, protrusion of the tongue, and inability to eat. Terminally, goats showed posterior paralysis, lateral recumbency, paddling, nystagmus, and pupillary dilatation. Many goats had food hanging from the mouth (Umoh 1977).
In a separate report of experimental caprine rabies involving two male goats 3-4 months of age infected with a fox-derived isolate in the masseter muscle, both goats initially showed an exacerbation of sexual behavior, priapism, and aggressiveness, followed progressively by incoordination, recumbency, opisthotonos, and myoclonia. The two goats died three and five days after onset of signs (Gomes et al. 2005).
Clinical Pathology and Necropsy
Laboratory confirmation of rabies in the living animal is difficult. Immunofluorescent staining of corneal scrapings or hair follicles in skin biopsies taken during clinical disease is confirmatory if positive, but false-negative results occur (Umoh and Blenden 1982). In one case of confirmed caprine rabies, CSF analysis was performed on a sample taken from the live animal. Analysis revealed a mononuclear pleocytosis, hyperproteinemia (46.7 mg/dL, normal range 24-40 mg/dL) and high glucose levels (146 mg/dL, normal range 45-87 mg/dL) (Moreira et al. 2018). Gross lesions are minimal at necropsy. There may be evidence of trauma caused by maniacal behavior. The meninges and brain may be congested and edematous. Histologically, neuronal degeneration with swelling and necrosis may be present throughout the brain, with a striking absence of inflammatory response. The most significant finding is Negri bodies in the cytoplasm of neurons, particularly in the hippocampus and in Purkinje cells. This finding has been considered diagnostic for rabies in the past, but falsepositive interpretations may occur and histologic techniques such as Seller staining for identification of Negri bodies are no longer recommended for diagnosis (OIE 2018d). Furthermore, Negri bodies may be absent or overlooked in cases confirmed by DFA testing or mouse inoculation (Silva et al. 2010).
Currently, immunofluorescence staining of brain sections is routinely carried out for rapid, accurate diagnosis of rabies, and the DFA test is the method of choice for rabies diagnosis. Impression smears of hippocampus, medulla, cerebellum, or gasserian ganglion are the preferred samples. Smears from brain stem, thalamus, and pons may also be diagnostic. If brain samples are submitted they should be fresh or glycerol preserved, because formalin fixation impairs performance of the DFA test. Peroxidase conjugate may be used on sections of formalin-fixed tissue for immunohistochemical tests in lieu of the DFA test. At a minimum, central vermis and right and left hemispheres of the cerebellum plus a complete cross-section of the brain stem should be submitted to the laboratory for analysis.
Not all countries have the laboratory capacity to run the DFA test because it requires expensive, specialized fluorescent microscopes, so alternative primary diagnostic test methods may be used, notably the direct rapid immunohistochemistry test (dRIT) or a pan-lyssavirus polymerase chain reaction (PCR) assay, of which there are two - the conventional reverse transcriptase PCR and the real-time reverse transcriptase polymerase chain reaction (RT-PCR) for viral RNA detection (OIE 2018d). For conducting epidemiologic surveys, an enzyme-linked immunosorbent assay (ELISA) that detects rabies antigen is recommended (Xu et al. 2007).
The most definitive, but cumbersome, diagnostic test is intracerebral inoculation of mice with brain, spinal cord, or salivary tissue of suspect cases. When positive, weanling mice develop rabies and virus can be identified by immunofluorescence in their brains. Cell culture in neuroblastoma cell lines with confirmation by fluorescent antibody testing is also available as an alternative to mouse inoculation. Mouse inoculation and cell culture should be used as confirmatory tests if DFA test, dRIT, other antigen detection tests, or PCR give inconclusive results (OIE 2018d).
Serologic tests are also available for detection of antibody to rabies, including virus neutralization (VN) and ELISA assays. They are used mainly for postvaccination surveillance in rabies control programs and for measuring antibody titers in individual animals.
Diagnosis
Regardless of the species of animal affected, rabies is one of the most difficult diseases to diagnose clinically because of the diversity of potential presentations. Veterinarians working in countries where rabies is enzootic should always consider rabies in the differential diagnosis when goats show any behavioral change or neurologic abnormality. Potentially, rabies can mimic all of the various neurologic diseases of goats. As has been so aptly stated, rabies is an unpredictable disease - the only characteristic feature is that it is uncharacteristic in its presentation (Rupprecht et al. 2002). Every reasonable attempt should be made to arrive at a definitive diagnosis other than rabies while the goat is alive. If a goat with signs of neurologic or behavioral dysfunction dies undiagnosed, it is incumbent upon the veterinarian to pursue a postmortem diagnosis of rabies.
Treatment and Control
There is no treatment for rabies. The zoonotic potential of rabies must always be considered when dealing with suspect cases. Such cases should be isolated and human contact kept to a minimum. Gloves and masks should be worn during examination or when performing necropsies, and hands washed thoroughly afterward with disinfectant soaps. When the suspicion of rabies is high, euthanasia should be recommended to expedite a definitive diagnosis at necropsy. A careful history of human contact should be documented and all people potentially at risk should be apprised of the situation and referred to physicians for postexposure counseling in the event that the suspect animal is confirmed as positive.
Based on incubation times reported in experimental caprine rabies, goats with suspected exposure should be strictly isolated and observed for at least one month in anticipation of the possible development of clinical rabies. Given the limited amount of information available on rabies in goats, it is possible that the incubation period for goats might exceed one month. Therefore, any goat showing clinical signs consistent with rabies during (or after) the observation period should be euthanized and submitted for necropsy diagnosis.
With regard to the potential benefit of postexposure prophylaxis (PEP) for goats exposed to rabies, the information available provides mixed guidance. In one study, repeated vaccination alone or with administration of antisera prolonged the incubation period for as long as 174 days in experimentally infected goats, but in no case did it prevent development of clinical disease (Umoh and Blenden 1981). A more recent report from Texas (Wilson et al. 2010) describes implementation of the Texas law that allows PEP in unvaccinated domestic animals exposed to rabies. The protocol requires immediate vaccination of the exposed animal, strict isolation of the animal for 90 days, and administration of booster vaccinations during the third and eighth weeks of the isolation period. Over a 10year interval, 1014 animals received PEP and no failures were recorded. While the majority were dogs and cats, some ruminants were involved, including four goats. Three levels of exposure were established, but the level of exposure for the goats was not stated. Vaccination of goats in enzootic areas may be possible according to local regulations. In the United States there is currently no vaccine specifically approved for use in goats, though several commercial killed rabies vaccines derived from cell cultures are approved in the country for use in cattle and sheep. Some veterinarians have been using killed vaccines approved in cattle and sheep in goats in the United States. The vaccines are administered initially to goats 3 months of age or older and they are boostered a year later. As there are no specific label recommendations for use in goats, subsequent revaccination schedules are empirical, with revaccination annually as in cattle or every three years as in sheep. Annual revaccination of goats is preferred, particularly under circumstances where there is a high risk of exposure or where increased human contact is expected, such as with goats at shows, fairs, and petting zoos. Also, as there is no goat-specific information, withholding goats from slaughter for a minimum of 21 days, as suggested for sheep post vaccination, is recommended.
Good client communication is essential when using rabies vaccine off label. Regulatory agencies in the United States do not recognize goats as officially vaccinated since the vaccine is not approved for that species. Therefore, in situations involving possible human exposure, vaccinated goats would be considered unvaccinated and in the United States could be subject to quarantine for up to six months or to euthanasia, depending on the situation and the associated regulatory requirements (Brown et al. 2016). Owners should be made aware of this possibility.
According to data from the Food and Agriculture Organization, rabies vaccination in goats is permitted in approximately 20 countries worldwide, including Brazil, India, Nepal, and the former Soviet Union (FAO 1989). The use of modified live rabies vaccines not approved for use in goats is strongly discouraged or prohibited.
Complete control of rabies depends on national and international efforts. Beyond vaccination, there is little that individual herd or flock owners can do to control rabies, except to know the likely reservoirs in their areas and minimize goat contact with them through housing, fencing, or animal control measures. Farm, herding, and other dogs in potential contact with goats should be vaccinated.
Pseudorabies
This acute fatal viral disease, also known as Aujesky's disease, infectious bulbar paralysis, or mad itch, is primarily a disease of swine and is reported only sporadically in goats. Clinical signs include peracute death or hyperexcitability, intense pruritus, convulsions, and terminal coma. Reported cases of caprine pseudorabies have invariably included direct or indirect contact with pigs.
Etiology
The causative agent is a double-stranded DNA, enveloped alphaherpesvirus. It is known by several names, including pseudorabies virus (PRV), porcine herpesvirus type 1 (PHV-1), and suid herpesvirus type 1 (SuHV-1). The virus is quite stable. It can survive in saliva, carcasses, and clean damp bedding for several months in winter, and several weeks in summer. Free, unprotected virus can persist for one to two weeks. However, the virus can be inactivated by a number of disinfectants, including sodium hypochlorite, formalin, peracetic acid, phenolics, and quaternary ammonium compounds.
Epidemiology
The virus is extremely well adapted to domestic swine, which serve as the agent reservoir. Adult pigs usually show only mild disease when infected, though abortions or early embryonic death can occur in pregnant sows. Affected piglets may show signs of neurologic disease and high mortality. Typical of the herpesviruses, latent infections and a carrier state occur in infected, recovered swine that can shed the virus in saliva and nasal discharges. All other susceptible species, including goats, cattle, sheep, dogs, and cats, are essentially dead-end hosts. They develop acute neurologic disease and die quickly.
In reported cases of caprine pseudorabies, the goats have been housed with or in fence contact with pigs (Herweijer and De Jonge 1977; Baker et al. 1982). The pigs may be latent carriers that shed virus, but show no evidence of infection. Other potential sources of exposure include transport vehicles in which infected swine have recently been hauled as well as modified live virus vaccines. Pseudorabies has occurred in sheep that were given parenteral medication using an uncleaned syringe previously used to vaccinate pigs for pseudorabies.
Transmission of virus is by inhalation or via skin abra - sions. Morbidity can be high, with 80% of exposed goats contracting the disease in an outbreak in the Netherlands (Herweijer and De Jonge 1977). The mortality rate is 100%.
Pathogenesis
The incubation period is one to four days. When the virus is introduced through the skin, it enters peripheral nerves and produces the characteristic localized pruritus that is observed in some cases of ruminant pseudorabies (“mad itch”). The virus then moves centripetally and enters the CNS, producing a rapidly fatal encephalitis or
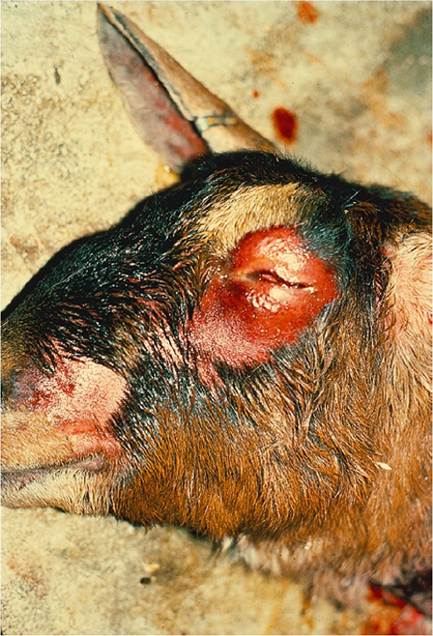
Figure 5.1 Periorbital alopecia and inflammation due to excessive rubbing of the face from pruritus due to pseudorabies infection in a goat having fence line contact with pigs. Source: Baker et al. 1982 / Rockwater.
myeloencephalitis. With inhalation of the virus, signs of encephalitis occur without a prodrome of pruritus.
Clinical Findings
All ages, breeds, and sexes can be affected. Goats may be found dead with no warning. Alternatively, they may show signs of intense rubbing or licking and even self-mutilation associated with severe, localized neurogenic pruritus (Figure 5.1). In cases where pruritus does not occur, initial signs may include agitation or excitation, repeated lying down and rising, hysterical bleating, profuse sweating, and convulsions. Fevers as high as 41.5 °C (106.7 °F) may be recorded. These excitatory signs give way to recumbency, paralysis with bloat and dyspnea, coma, and finally death. The clinical course may be only several hours up to 24 hours.
Clinical Pathology and Necropsy
While serology is useful in the identification of infected swine, it has no application in caprine pseudorabies because goats become clinically ill and die before an antibody response can develop. At time of necropsy, attempts should be made to isolate the virus from brain tissue. There are no obvious gross necropsy findings, but there are histologic lesions of severe, focal, non-suppurative encephalitis and myelitis. Eosinophilic intranuclear inclusion bodies typical of herpesviruses may be found in degenerating neurons. Immunofluorescence and immunoperoxidase methods can be used to confirm the identity of the SuHV-1 in infected brain tissue. A PCR technique for detecting the virus genome in mixed tissue homogenate samples from the field has also been described (Perez and de Arce 2009).
Diagnosis
When pruritus is present, primary dermatologic conditions, particularly ectoparasites, must be ruled out. Scrapie, rabies, and cerebrospinal nematodiasis can also produce neurogenic pruritus in goats. When encephalitic signs predominate, consider also rabies, cowdriosis (heartwater), PEM, and hypomagnesemia. The acute nature of the disease also suggests poisonings, notably cyanide, nitrates, urea, organophosphates, and chlorinated hydrocarbons.
Treatment and Control
There is no treatment for pseudorabies. Control depends on recognizing the risks of commingling goats with pigs whose pseudorabies status is unknown. If pigs must be housed with goats, the pigs should be serologically negative for pseudorabies before purchase. National regulatory programs to eradicate pseudorabies in commercial swine greatly reduce the risk of goats contracting the disease. In the United States, a pseudorabies eradication program was initiated in 1989, and by 2004 all 50 states were declared free of pseudorabies in commercial swine. However, feral swine serve as a reservoir of pseudorabies infection in the United States, particularly in the southeastern and southcentral states (Pedersen et al. 2013), and while commercial swine in total confinement continue to remain free, there has been spillover from feral swine into domestic swine managed with access to the outdoors. Outdoor goats too could be exposed in this way.
Historically, pseudorabies has not been considered a public health risk. However, there have been some putative reported cases of pseudorabies from Europe in people exposed to cats. Antibodies to PRV were documented in these individuals (Mravak et al. 1987). More recent reports from China regarding human cases of encephalitis associated with PRV suggest that pseudorabies could be an occupational risk for pig farmers and pork industry workers (Wong et al. 2019). Therefore, veterinarians and others working directly with the virus or with pseudorabies- infected animals or carcasses should observe appropriate biosecurity and biosafety precautions.
Scrapie
Scrapie is an infectious, contagious, degenerative neurologic disease that occurs naturally only in sheep and goats. It belongs to the group of diseases known as transmissible spongiform encephalopathies (TSEs), which are caused by abnormal configurations of naturally occurring proteins known as prions. Prions are unique as infectious agents in that they contain no genetic material. Other TSEs include bovine spongiform encephalopathy (BSE) of cattle; chronic wasting disease of deer and elk (CWD); transmissible mink encephalopathy of farmed mink (TME); and several diseases of humans, including Kuru, Creutzfeldt-Jakob disease (CJD), and variant Creutzfeldt-Jakob disease (vCJD).
Scrapie is characterized by an extended incubation period of months to years. When clinical signs ultimately develop, they may include weight loss, incoordination, changes in mental status, progressive debilitation, and pruritus. Historically, clinical reports of scrapie in goats have been less common than in sheep, but the goat industry has become more focused on the disease as goats are now regularly included in regulatory surveillance and control programs for TSEs, including scrapie, and the occurrence of the disease in goats has become clearer. Veterinarians, and the goat owners they serve, must be aware of the details of these control programs, because they can affect the management and commercial aspects of goat production.
Interest in and knowledge of scrapie notably increased as a result of the emergence of BSE, a TSE that is transmissible to humans as vCJD (Bruce et al. 1997). BSE has numerous clinicoepidemiologic similarities to scrapie and this has sparked concerns that BSE could also occur in small ruminants and therefore needs to be differentiated from scrapie. As a result, better tools are now available for the diagnosis of scrapie, there is a clearer understanding of the pathogenesis, and, because of active surveillance programs, there is also more epidemiologic knowledge about scrapie in goats. Despite similarities to BSE in cattle, which is zoonotic, scrapie of sheep and goats is not believed to be transmissible to humans under natural conditions (EFSA 2015).
Etiology
The cause of scrapie and other TSEs in animals is the expression of the abnormal form of a naturally occurring cellular protein called PrPC. This protein can be found in cells throughout the body, but is of clinical and diagnostic interest primarily in the nervous system and the lymphore- ticular system (LRS). It is a membrane glycoprotein bound to the outer surface of neurons, lymphocytes, and other cells. The most well-known activity of normal cellular PrPC is its ability to bind copper and other metal ions through its flexible N-terminal moiety, though the exact metabolic functions associated with this metal-binding ability are not fully elucidated (Salzano et al. 2019). The role of PrPC in copper metabolism, neuroprotection, synaptic transmission, neuronal excitability, and other functions continues to be explored (Wulf et al. 2017). The structure and patho- biology of prions and the diseases they cause in animals and humans have been reviewed (Prusiner 1998; Aguzzi and Heikenwalder 2006; Aguzzi 2006; Greenlee and Greenlee 2015; Zabel and Reid 2015).
Through mechanisms that are not fully understood, the development of scrapie is associated with abnormal replication of normal host prion protein PrPC to produce a prion protein of modified structure, PrPSc (also referred to in the literature as disease-associated protein, PrPd), the accumulation of which occurs simultaneously with degeneration of nervous tissue and other manifestations of disease. This process is complex, with several factors involved: exposure of the animal to abnormal forms of PrPSc that induce abnormal replication of PrPC in the host; variations in the strain of PrPSc encountered; and variations in the genetic makeup of the animals exposed to PrPSc. Together these factors can influence host susceptibility to the production of abnormal endogenous PrPSc following exposure, the incubation period for manifestation of disease, and the severity of disease. The details of these interactions remain topics of active research.
PrPC protein is composed of 210 amino acids. Structurally, it contains a highly flexible N-terminal domain and a structured C-terminal domain consisting of three helices with α-helical folding and a short β-sheet. The molecule contains two N-glycosylation sites and the naturally occurring protein may comprise mono-, di-, or non-glycosylated forms. The fully glycosylated form has a molecular weight of 33-35 kDa. The ratio of the three forms in tissue extracts of PrPsc is referred to as the glycotype. Differences in patterns of glycosylation are one tool used to differentiate various strains of the scrapie prion.
In scrapie, as in most other TSEs, the key factor in the pathogenesis of the disease is the conversion of normal PrPC protein into an isoform, in this case PrPsc, which is characterized by insolubility and partial resistance to degradation by proteases. Sometimes the term PrPres may be used in the literature to refer to this resistance property of PrPSc, while PrPC may be referred to as PrPsen to indicate its sensitivity to proteolytic degradation. The isoform demonstrates important secondary structural change compared to the normal PrPC protein. This is manifested as aberrant folding of the protein molecule that results from one of the three normal α-helices being replaced by a β- sheet or β-helix structure. The conversion from PrPC to PrPSc has been described as a stabilization of a proto-β-helical motif by a neighboring PrPSc molecule and subsequent extension to form the complete β-helix (Wille et al. 2002). Different types of the scrapie prion protein exist and they are characterized either by their behavior in hosts (e.g., incubation periods, clinical manifestations, and histologic distribution of lesions) or by the physical characteristics of the prion protein itself (e.g., structural, biochemical, and stability differences). In sheep over 20 different strains of scrapie have been identified, some experimentally after serial passage in mice, and the natural occurrence in sheep of some of these strains remains undetermined. In goats specifically, knowledge on prion strain characterization is limited. In a recent study of TSE- infected goat brains identified through TSE surveillance in seven European countries, two different scrapie types were found: Nor98∕atypical scrapie and classical scrapie. However, classical scrapie was associated with three distinct PrPSc variants, CS-1, CS-2, and CH1641, distinguishable by the protease sensitivity of the N-terminus of PrPSc (Langeveld et al. 2019).
The atypical type of scrapie was first reported from Norway in association with five clinical cases of atypical sheep scrapie with unusual clinical and pathologic features, the first of which was seen in 1998. The strain that is responsible for atypical scrapie has been designated as Nor98. Among its unusual features are the fact that it produced disease in sheep with genotypes ordinarily associated with resistance to scrapie; an atypical distribution of lesions in the brain, with a notable absence of lesions at the level of the obex; the absence of detectable PrPSc in lymphoid tissues using immunohistochemistry (IHC) and ELISA; and a distinctive molecular PrPSc profile on western blot assay, which indicates a glycotype that is distinct from classical scrapie and the prion strain producing BSE (Benestad et al. 2003). In 2007, the first sheep in the United States, originating from Wyoming, was confirmed with the Nor98 type of scrapie through slaughter surveillance (USDA 2007a).
Scrapie prions are very resistant to physical and chemical destruction and are believed to persist on pastures for months to years when excreted by infected sheep or goats. Standard disinfectants do not destroy prions. Recommended disinfection procedures include deactivation with 2% sodium hydroxide solution, steam sterilization at 132 °C for one hour, or incineration. For decontamination of premises, four applications of sodium hypochlorite solution (20 000 parts per million [ppm] free chlorine) one hour apart have been evaluated (Gough et al. 2017), but in one study scrapie prion-contaminated farm premises thought to be effectively decontaminated were shown to be rapidly recontaminated, presumably through the persistence of prions in dust (Gough et al. 2019).
Epidemiology
Most nations where small ruminants are raised have never reported scrapie to the World Organisation for Animal Health (OIE), the intergovernmental agency that monitors global animal disease status. However, it is unclear whether these countries are actually free of the disease, or if the disease exists undetected or unreported due to the lack of active surveillance programs and a lack of recognition by farmers of the clinical picture of scrapie.
Scrapie is enzootic in numerous countries of Europe, including the United Kingdom, as well as in Iceland, the United States, and Canada. It also has been reported in Australia, New Zealand, India, South Africa, Kenya, Brazil, and Colombia as a result of sheep importations from the United Kingdom from the 1930s through the 1970s. Australia and New Zealand eliminated the disease in the 1950s and have remained free through surveillance and strict importation rules. South Africa eliminated the disease in 1972 and also remains free.
Surveillance activities for TSEs in small ruminants have revealed that atypical scrapie may also occur in countries where classical scrapie is present, but at much lower rates. Surveillance has also revealed that atypical scrapie is found in countries where classical scrapie is not present. A case of atypical scrapie/Nor98 was identified in a New Zealand sheep in 2009 (Kittelberger et al. 2011) and in several sheep in Australia (Cook et al. 2016). In Europe, atypical scrapie has been identified in sheep in Portugal, Germany, and Austria over a 14-year period, though no classical scrapie was detected in those three countries during the same period (Ricci et al. 2017). These findings underscore that atypical scrapie is a condition distinct and separate from classical scrapie.
Spurred by the emergence of BSE in cattle in Europe and the subsequent association of BSE with vCJD in humans, the European Union (EU) initiated aggressive surveillance activities for TSEs in small ruminants. From 2002 to 2005 inclusive, for all 25 EU member countries, a total of 420 299 goats were tested for scrapie and 1669 (0.4%) were positive. These goats included healthy animals at slaughter as well as animals dead on farm and suspect clinical cases. In comparison, 1 511 375 sheep were tested during this same period and 8930 (0.6%) were positive (European Commission 2006b). This indicated that the prevalence of scrapie infection in the goat population is not that different from the sheep population.
Active small ruminant surveillance for TSEs in the EU is ongoing. Between 2006 and 2018, approximately 2.05 million goats from within the then 28 EU member states plus four additional non-member European countries were screened for TSEs at slaughter or post mortem. During that period, 12 542 classical scrapie cases were identified in goats, as were 163 atypical scrapie cases, a prevalence of approximately 0.6% for classical scrapie and 0.01% for atypical scrapie (EFSA 2019). The first case of atypical scrapie in a goat outside of Europe was reported from Japan in 2019 (Matsurra et al. 2019).
Historically, reports of naturally occurring clinical scrapie in goats have been less common than in sheep. It is not clear if this was due to a comparatively lower incidence or a failure to recognize the disease in goats. Naturally occurring caprine scrapie was reported in France in 1942 (Chelle 1942), the United States in 1969 (Hourrigan et al. 1969), Canada in 1975 (Stemshorn 1975), Switzerland in 1982 (Fankhauser et al. 1982), Cyprus in 1989 (Toumazos and Alley 1989), and the United Kingdom on several occasions through the 1960s and 1970s (MacKay and Smith 1961; Brotherston et al. 1968; Harcourt and Anderson 1974). Clinical caprine scrapie also has been reported from Italy in 1998 (Capucchio et al. 1998), Greece in 2002 (Billinis et al. 2002), and Finland in 2002 (Government of Finland, 2002).
In the United States from 1990 to 2007, there were 19 cases of caprine scrapie confirmed by regulatory authorities. The distribution of these cases was five in California; three in Colorado; two each in Washington, South Dakota, and Ohio; and one each in Wyoming, Nebraska, Illinois, Michigan, and New Hampshire (USDA 2007b).
From 2001 to 2003, the United States Department of Agriculture (USDA) conducted the Scrapie Ovine Slaughter Surveillance Study (SOSS) on healthy, mature market sheep to determine the national prevalence of the disease, which was determined to be 0.2% (USDA 2004). At the end of fiscal year (FY) 2016, the percent of cull sheep found positive at slaughter was 0.001% with an upper confidence limit of 0.009%, representing a 99% decrease since slaughter surveillance started in FY 2003. Goats were not included in the 2001-2003 national study. However, USDA conducted a study from May 2007 through March 2008 to estimate the prevalence of scrapie in goats in the United States. The study featured goats with a potentially higher likelihood of infection combined with routine samples collected through Regulatory Scrapie Slaughter Surveillance (RSSS), and the prevalence of classical scrapie in goats was determined to be less than 0.1%. The national prevalence in goats was estimated to be below 0.02% from FY 2016-2018 (USDA 2019).
Evidence to date indicates that atypical scrapie is not a transmissible disease, but rather occurs spontaneously in older goats and sheep (Greenlee 2019). Classical scrapie is certainly transmissible and transmission in sheep is mainly horizontal, from dams to lambs through contact with contaminated placentas post lambing, but also directly from animal to animal or though introduction of susceptible sheep into environments contaminated with the scrapie prion. Vertical transmission via milk and colostrum from ewe to lamb has been demonstrated (Konold et al. 2013), as has transmission of scrapie to lambs fed milk from scrapie-infected goats (Konold et al. 2016). The role of other excretions or body fluids in the transmission of scrapie remains unclear. Newer amplification techniques for detection of misfolded prion proteins have now detected them in plasma, urine, and saliva of sheep. Progress in this area has been reviewed (Properzi and Pocchiari 2013).
Vertical transmission in utero is not considered to play a significant role in the epidemiology of scrapie, but the presence of scrapie prions in fetal lamb tissues has been documented (Spiropoulos et al. 2014), as has evidence of prenatal transmission from ewes to lambs in genotypically scrapie-susceptible sheep (Foster et al. 2013). It is also considered possible for scrapie to be transmitted through embryo transfer, but that risk is significantly reduced if the embryos are derived from sheep with ARR scrapie-resistant genotypes (EFSA 2010) and the transfer is conducted according to the standards set for embryo transfer by the OIE (OIE 2021a). Whether or not scrapie is transmissible through semen remains unclear. The European Food Safety Authority (EFSA) issued a scientific opinion in 2010 that the risk of TSE transmission associated with sheep and goat semen ranges from negligible to low, but noted that the information available for goats was less robust than for sheep (EFSA 2010). Nevertheless, in 2012 there was a report from the United States involving sheep research flocks which indicated that scrapie was transmitted through breeding from a scrapie-positive ram to a genotypically susceptible ewe, with evidence of infectivity of ram semen confirmed by amplification procedures (Rubenstein et al. 2012). The evidence for and against the role of prenatal transmission of scrapie in sheep and goats and its significance for veterinary public health have been reviewed (Adams 2016).
In the majority of caprine cases occurring worldwide, there is a history of contact between affected goats and infected sheep, though direct contact with sheep is not an absolute prerequisite for caprine infection. In the Canadian report, five years had elapsed between the last case of ovine scrapie on a property and the first case in goats, although sheep remained on the farm (Stemshorn 1975). In one report from the UK, there was no discernible contact of four affected Saanen goats with scrapie-infected sheep (Harcourt and Anderson 1974). Wood et al. (1992) reported on 20 caprine scrapie cases seen at the UK Central Veterinary Laboratory since 1975. In at least seven of those cases, it was established that there had been no direct or indirect contact with sheep.
The principal method of natural transmission from sheep to goats is horizontal via direct contact with infected placentas or by indirect contact with pastures or bedding contaminated by such placentas (Pattison et al. 1972). The presence of the infective agent PrPSc has been confirmed in the placentas of infected sheep (Tuo et al. 2001) and in the placentas of naturally infected goats, though the levels of PrPSc in goat placentas were lower than generally found in sheep placentas (O'Rourke et al. 2011). The presence of infectivity in milk or colostrum, as mentioned earlier, would contribute to infection of lambs or kids.
Iatrogenic transmission is also possible. Outbreaks of scrapie involving goats in Italy between 1997 and 1999 were linked to vaccination of goat and sheep herds with a Mycoplasma agalactiae vaccine for contagious agalactia. The vaccine had been prepared from ovine CNS, lymph node, and mammary tissues that apparently contained the scrapie prion (Agrimi et al. 1999; Caramelli et al. 2001). An earlier outbreak of scrapie in sheep in the United Kingdom in the 1930s was associated with the administration of a vaccine for louping-ill prepared from sheep nervous tissue apparently contaminated with the scrapie prion (Gordon 1946). In a more recent report, the detection of PrPSc in only the prescapular lymph node of a goat from a known infected herd raised the possibility of iatrogenic infection due to subcutaneous (SC) injection with a contaminated needle in the neck region (Konold et al. 2020).
Studies from Iceland suggest that iron and manganese content of soils and forages may influence the occurrence of scrapie in that country. Forages from scrapie-afflicted farms had significantly higher iron concentrations and iron/manganese ratios than forages from scrapie-free farms. It was speculated that manganese may in some manner inhibit absorption of prions from the gut, and that high iron levels suppress manganese in the diet and thereby facilitate the absorption of prions (Gudmunsdottir et al. 2006).
Pathogenesis
Experimentally, the disease has been produced in goats by SC and intracerebral inoculation of the agent and by feeding of fetal membranes from infected sheep (Pattison 1957; Pattison et al. 1972; Hadlow et al. 1974). In naturally occurring scrapie, it is most likely that oral ingestion of the infective prion initiates new infections. Following ingestion, the portal of entry for the scrapie agent may be the Peyer's patches of the ileum (Heggebo et al. 2000), from which it is transported via blood or lymphatics to other sites in the LRS, including tonsil, spleen, and retropharyngeal and mesenteric lymph nodes. Presence in the gut-associated lymphoid tissues likely facilitates access of the infective agent to the enteric autonomic nerves, which may in turn provide access to the CNS, though the precise mechanisms for neuroinvasion in the CNS remain an area of active research. Replication in the LRS continues for weeks to months before infectivity can be detected in the brain, and it may be years before there is clinical manifestation of disease.
In experimentally infected goats, inoculated intracerebrally or SC, the infective agent is disseminated to a variety of tissues in different levels of concentration, based on subsequent mouse inoculation assays conducted with titrations of various tissues (Hadlow et al. 1974). Brain and spinal cord had the highest levels of infectivity. Moderate levels were found in retropharyngeal, superficial cervical and subiliac lymph nodes, spleen, tonsil, and adrenal gland. Low levels were found in CSF, sciatic nerve, pituitary gland, nasal mucosa, ileum, proximal colon, distal colon, liver, thymus, mediastinal and bronchial lymph nodes, and parotid salivary gland. No infectivity was found in blood clot, submaxillary salivary gland, thyroid, heart, lung, kidney, skeletal muscle, bone marrow, pancreas, ovary, and saliva.
Once the infective agent has reached the brain, it induces PrPC, the normal cellular form of PrP, to undergo a conformational change, resulting in increased β-sheet folding and subsequent appearance of PrPSc, often as scrapie-associated fibrils in the nervous tissue. The exact mechanisms of this process at the molecular level are currently an area of active research. The net result of this protein misfolding and fibril accumulation is a progressive degeneration of nervous tissue characterized by a sponge-like pattern of vacuolation, which leads to the general description of scrapie and other TSEs as spongiform encephalopathies. The brain lesions of scrapie are described further in the section on necropsy findings.
Susceptibility to disease and the incubation period are now known to be genetically controlled in sheep and goats (Baylis and Goldmann 2004), though at present the relationships are more clearly defined in sheep than in goats. The mammalian PRNP gene, depending on species, is composed of two or three exons with the entire open reading frame (ORF) contained in the last exon. The gene that codes for PrP protein in goats and sheep has an ORF of 256 codons in length, with posttranslational processing resulting in a mature PrPC protein of 210 amino acids. Polymorphisms occur in the protein coding region of the PRNP gene that are known to be associated with susceptibility to scrapie and the incubation period of the disease in sheep. Such associations also occur in goats, and more has become known about them in recent years.
At present, three specific ovine PrP polymorphisms are definitively known to influence the susceptibility for development of the disease in exposed sheep. These occur at codons 136, 154, and 171 and are expressed as A136V, R154H, and Q171R/H, respectively. The amino acid valine (V) instead of alanine (A) is encoded at 136, histidine (H) instead of arginine (R) at codon 154, and R or H instead of glutamine (Q) is encoded at position 171. These are usually written in shorthand, so that, for example, a sheep with the homozygous genotype A136, R154, R171/A136, R154, R171 would be referred to as the ARR/ARR genotype. Notably, the polymorphisms at codons 136, 154, and 171 that are associated with increased susceptibility to classic scrapie in sheep have no bearing on the disease in goats. However, in both sheep and goats, H154 is associated with an increased susceptibility to atypical scrapie (Colussi et al. 2008).
By the year 2000, the relationship of specific polymorphisms with resistance or susceptibility to classical scrapie in sheep was sufficiently well established that selective breeding programs based on identifiable polymorphisms were being integrated into national scrapie control and flock certification programs and, based on level of compliance, they have proven effective in reducing the prevalence of the disease in sheep (EFSA 2014). This success has spurred considerable interest in determining whether or not such polymorphisms related to scrapie susceptibility or resistance exist in goats.
Between 1995 and 2006, multiple research investigations identified numerous PrP polymorphisms in goats of different breeds, but their relationship to scrapie susceptibility was not always clear. Bayliss and Goldmann (2004) identified many polymorphisms, including W102G, T110P, G127S, I142M, H143R, R154H, P168Q, R211Q, Q220H, and Q222K. Association with disease was suggested for three of them, namely I142M, H143R, and R154H, and also possibly for P168Q and Q222K. Polymorphism I142M in particular is associated with differing scrapie incubation periods in response to various prion strains (Goldmann et al. 1996).
Additional PrP polymorphisms have been reconfirmed in different breeds. Billinis et al. (2002) reported such polymorphisms for V21A, L23P, and G49S in goats in Greece. For the Ionica breed of goats in Italy, Vaccari et al. (2006) reported polymorphisms for G37V, T110P, H143R, R154H, Q222K, and P240S, with the suggestion that the R/H polymorphism at codon 154 does not confer scrapie resistance, but does plays a role in the elongation of scrapie incubation time in infected goats, while the presence of lysine (K) at codon 222 is associated with resistance to classical scrapie.
Two additional polymorphisms not previously reported in goats were identified in Italian goats by Acutis et al. (2006), namely L133Q and M137I. In addition, the authors also noted a protective role against scrapie in goats for the glutamine to lysine mutation at codon 222 (Q222K). Zlhang et al. (2004) evaluated five Chinese breeds of goats for PrP polymorphisms. All breeds possessed the R154H polymorphism and the authors also identified a previously unreported polymorphism, I218L, in two of the breeds. By 2008 PRNP gene polymorphisms in codons 21, 24,37,49,102,110, 127, 133, 137, 142, 143, 146, 151, 154, 168, 211, 218, 220, 222, and 240 had been documented in goats (White et al. 2008).
By 2017, sufficient information had become available on the relation of genetic polymorphisms to scrapie susceptibility in goats that a review of the existing data was commissioned by the EFSA to produce a scientific opinion on the role of genetics in resistance to classical scrapie and other TSEs (Ricci et al. 2017). The review concluded that the evidence for the role of K222 in genetic resistance to classic scrapie in goats was actually stronger than the evidence that existed in 2001 for the role of the ARR allele in sheep. In addition, it was confirmed that alleles D146 and S146 also confer genetic resistance, though the cumulative evidence is less strong. Evidence regarding nine different alleles was assessed overall and their relative strengths of resistance for classical scrapie were reported as follows:
K222 > D146 = S146 > Q211 = H154 = M142 > S127 = H143 > wild type.
Based on this analysis, it was suggested that breeding for resistance can be an effective tool for controlling classical scrapie in goats and that it could be an option at herd and population levels. However, the report included some caveats. It was noted that the alleles associated with resistance are heterogeneously distributed across the EU member states and goat breeds, often at low frequencies (to scrapie have the genetic polymorphisms H154, M142, and P240, where H represents histidine, M methionine, and P proline.
• Goats that are highly susceptible to scrapie are associated with the genetic polymorphisms Q222, R211, R154, N146, and I142, where Q represents glutamine, R arginine, N asparagine, and I Isoleucine.
Genetic resistance may not be absolute, particularly if the protective allele is heterozygous. Nevertheless, goats singly heterozygous for PRNP S146 or K222 that were orally inoculated with classical scrapie at birth show no disease at ages well beyond 6 years (Cinar et al. 2018). In a study in Cyprus, goats 6 years of age or older that were culled from naturally infected herds showed no evidence of scrapie if they were either heterozygous or homozygous for alleles D146 or S146 (Georgiadou et al. 2017). Experimental data supports the surveillance observations regarding the significance of polymorphisms at codon 146 on resistance to scrapie (Papasavva-Stylianou 2017).
Clinical Signs
Scrapie is a neurologic disease and clinical evaluation of goats suspected of scrapie should include careful assessment in three areas: changes of mental status, behavior, and activity; changes in sensory responses; and changes in movement, (Konold et al. 2010; Konold and Vallino- Costassa 2018). While no particular sign is pathognomonic for scrapie, any clinical signs attributable to neurologic dysfunction in an adult goat, especially if seen in association with evidence of pruritus, should prompt consideration of a diagnosis of scrapie. Possible clinical signs associated with classical scrapie are summarized in Table 5.4. In a study designed to monitor clinical signs of TSEs in goats, it was noted that scrapie can present as clinical disease with or without pruritus; and that certain signs, notably tremor, positive scratch test, extensive hair loss, ataxia, and absent menace response, were highly disease specific for scrapie (Konold et al. 2010). In both sheep and goats it has been suggested that the presence of two or more of the following signs in an individual animal is highly suggestive for scrapie: a repeatable response to scratching; tremor; abnormal behavior; circling; collapsing episodes; ataxia; or an absent menace response (Konold and Phelan 2014). However, if goats are assessed for scrapie on clinical signs alone, many cases will be missed (Konold et al. 2020).
While goats most likely become infected with scrapie as neonates, clinical manifestations of the disease are not seen until adulthood, usually in goats at least 2 years of age and older. The clinical presentation may vary considerably, particularly in the early stages, and the clinical course may slowly progress over one to six months. The onset of signs
Table 5.4 Signs of classical scrapie that might be observed in individual goats, understanding that the presence ofspecific signs is highly variable.
Pruritis/skin irritation
Repeated rubbing of the head and body against objects
Repeated scratching of the flanks with feet or horns
Nibbling (lip licking or smacking) or grinding teeth when rubbing themselves
Nibbling, lip movements, and/or head movements when rubbed by person firmly on the back (positive scratch test)
Continued scratching of the shoulder or ear with a hind foot Excessive hair loss
Damage to the skin such as abrasions, excoriation, discoloration, hyperpigmentation, crusts, lichenification, and secondary bacterial infections.
Changes in behavior
Hyperalert and easily excitable if handled or approached
Ears pricked and/or tail cocked forward
Increased nervousness or exaggerated menace response
Running into objects (visual impairment)
Aggression
Lagging behind or separation from the herd
Depression or vacant stare
Resists milking
Changes in posture and movement
Trembling (mainly of the head)
Awkward stances, notably carrying the hindlimbs forward so that the rump is elevated and the withers are held low Unusual high-stepping gait
Circling when blindfolded (uncommon)
Stumbling
Collapsing when chased or handled
Severe incoordination
Weak hindlimbs
Difficulty standing
Recumbency
Other possible signs
Weight loss
Anorexia
Hypersalivation, slobbering
Difficulty swallowing
Regurgitation of rumen content
Premature cessation of milk production
Unexpected death (before recognition or appearance of other clinical signs)
is insidious. Goat and sheep farmers experienced with scrapie in their flocks may notice early non-specific signs such as weight loss or separating from the flock, which might be overlooked by others not familiar with the disease. Other early signs may include a loss of inquisitiveness and the development of an irritable disposition, which often first manifests as a resistance in lactating does to being milked.
In experimental infection of goats (Pattison et al. 1959), affected goats often showed a characteristic posture of carrying the rear limbs forward so that the rump was elevated and the withers were held low. In addition, the tail was held up, cocked forward over the rump, with the ears frequently pricked forward, suggesting heightened alertness. Restlessness increased and goats became hypersensitive to handling, with affected goats attempting to flee, bleat uncontrollably, or stiffen with muscular rigidity when handled. Left alone, they would hold the head down and stomp as if bothered by imaginary flies. Fine tremors sometimes occurred. Over time, behaviors of heightened alertness may give way to drowsiness or seeming drunkenness. Incoordination may become more pronounced, with increased stumbling and falling and increased difficulty rising. Pruritus is a prominent and often dramatic sign in sheep; the name of the disease derives from the historical observation of affected sheep scraping themselves raw on inanimate objects in response to the intense itching they experienced.
Pruritus can also be an important sign of scrapie in goats, but occurs less consistently and may be less severe than observed in sheep. Some goats may show localized pruritus, particularly over the withers or tail head. Nibbling at the skin rather than rubbing against stationary objects may be more likely in goats than in sheep. Horned goats may lean their heads back and scratch this area persistently with the horn tips or scratch, dog-like, with their hindlimb if the itchy spot is reachable. Administering a “scratch test” can assist with the diagnosis. While it is not always present in animals with clinical scrapie, scratching the suspect animal on its dorsum should produce characteristic elevated head movements and lip movements, the “nibbling reflex.”
Other, less common signs may include teeth grinding, salivation or slobbering, regurgitation of rumen contents, and impaired vision. Weight loss may become marked and anorexia develops in the terminal stages (Figure 5.2). Affected animals eventually become prostrate and die if not euthanized first. Fever does not occur in the course of the disease.
Clinicians and goat owners should keep in mind that the clinical presentation in individual goats may vary considerably. One confirmed case of scrapie in a goat showed no signs other than listlessness, progressive weight loss, and premature cessation of milk production (Harcourt and Anderson 1974). In another confirmed case, the only apparent clinical sign was regurgitation of rumen contents (Wood et al. 1992).
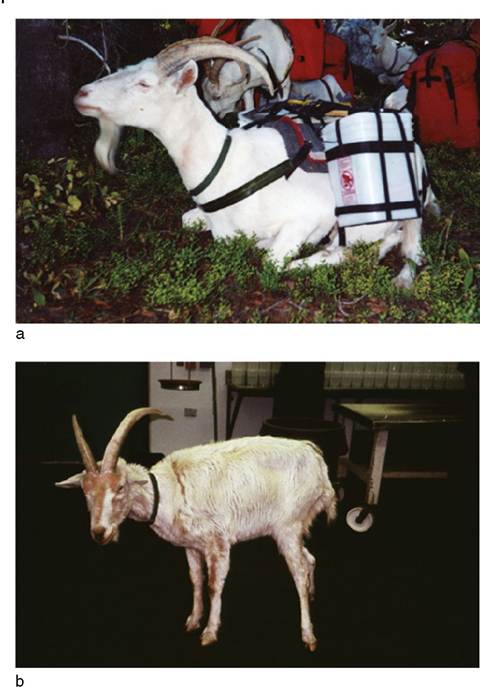
Figure 5.2 Clinical progression of a laboratory-confirmed case of scrapie in a Saanen wether goat. (a) The goat robust and healthy prior to the onset of clinical signs, and actively working as a pack goat. (b) The same goat in an advanced state of debilitation and emaciation 5 months later at 71 months of age. In addition to the apparent weight loss, the goat developed fearful behavior, difficulty swallowing, and pruritus, with hair loss over the face and forequarters. The goat herd from which this animal came had no history of association or contact with sheep. Source: Courtesy of Dr. David M. Sherman.
There are no consistent hematologic, biochemical, or gross pathologic changes in animals with clinical or pre- clinical scrapie that are diagnostic of the disease. However, antemortem diagnosis of scrapie may be possible by histologic examination of biopsy material from specific sites, as discussed further below.
Atypical scrapie is, for the most part, clinically indistinguishable from classical scrapie, both presenting with an overlapping array of similar neurologic signs. In fact, the vast majority of atypical scrapie cases have been diagnosed at slaughter or post mortem through TSE surveillance programs. Nevertheless, some aspects of the clinical presentation might suggest atypical scrapie. Atypical scrapie can occur in herds with no history or previous diagnosis of classical scrapie and it tends to affect older animals, 5 years of age or greater. In two goat cases confirmed at slaughter, the history included blindness, stiff gait, and apathy in one, while the other was just found dead (Nentwig et al. 2007).
Clinical Pathology and Necropsy
Scrapie has been known as a disease of small ruminants for more than 250 years, yet it is only in recent decades that new laboratory tests have become available for reliable confirmation of the disease. Early obstacles were a lack of understanding about the nature of the causative agent and the inability to culture it in vitro or identify it in vivo. Another important impediment has been that prion infection produces no detectable host humoral immune response, so that no serologic tests for detection of antibodies have been possible. While serologic tests remain unavailable, scientists now are able to produce monoclonal antibodies to specific epitopes of the PrPSc protein, and this has become a tremendously useful tool for better characterizing the scrapie agent and for identifying PrPSc in tissues through the use of immunobiologic tests such as IHC and electrophoresis followed by immunoblotting. The diagnostic procedures currently in use for the diagnosis of scrapie are summarized elsewhere (OIE 2018e).
Before the application of these techniques, diagnosis of scrapie was limited to histologic examination of affected brain tissue to identify characteristic lesions and confirmation of infectivity by inoculation of mice with tissues taken from suspect cases, a cumbersome procedure that can take up to two years to yield results.
Commercial immunoassays based on the ELISA format are used as rapid screening tests in TSE surveillance programs to initially detect the presence of abnormal prion protein in tissues because they can be automated, resulting in high output, quick turnaround times, and reduced financial costs. They were initially approved for use in screening cattle for BSE in 1999, but in 2002 the EU expanded their use to include screening of goats and sheep for scrapie. Subsequently, some of these tests have been withdrawn from the market due to reduced demand, since the BSE crisis is under control. Tests developed specifically for small ruminants have also been produced and authorized for use in the EU. However, these tests were assessed for sheep, and approval extrapolated for use in “small ruminants,” without taking into account that goats are a different species. As a result, the sensitivity of these tests has been questioned (Konold et al. 2020; Simmons et al. 2020) and it has been proposed that TSE tests should always be assessed specifically for the populations in which they are to be applied. The sensitivity of western blot and IHC to detect TSEs in goats is not affected (Simmons et al. 2020).
For more discriminatory testing, IHC techniques that use monoclonal antibodies to different PrPSc epitopes are available to detect the presence of PrPSc in formalin-fixed tissues. IHC techniques can be applied to biopsy materials taken from live animals or from sections of tissue obtained at necropsy.
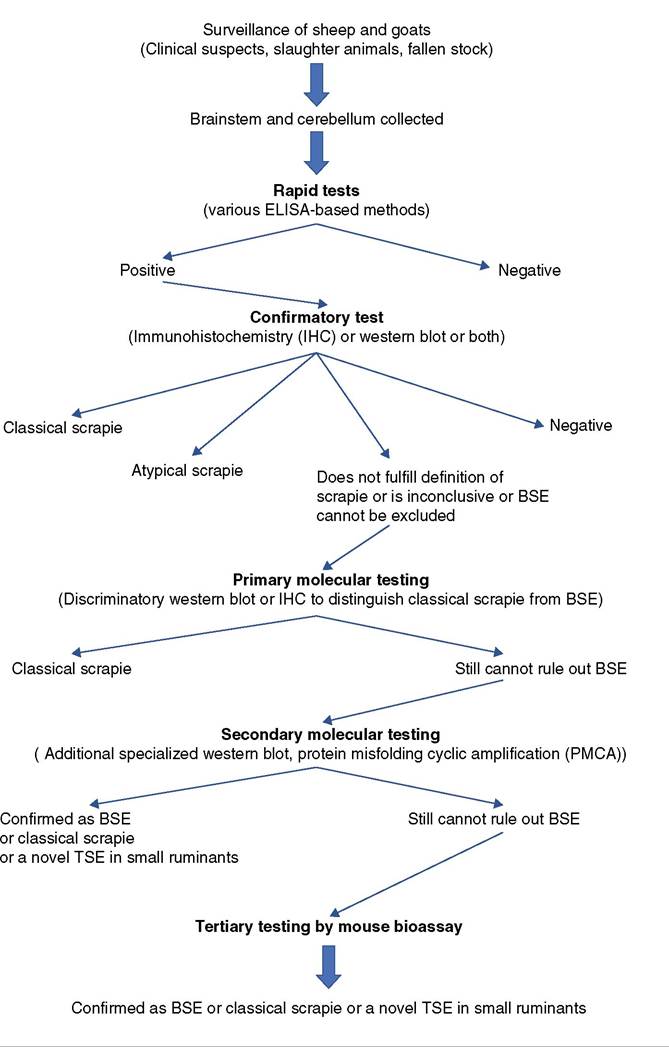
Immunoblotting by the western blot technique is performed on fresh tissues. It is used in regulatory surveillance and monitoring programs to screen for TSEs, including scrapie. Reference laboratories use western blot to distinguish different strains of scrapie prion, including atypical scrapie, and to differentiate PrPsc from that of the prion causing BSE. A flowchart for current testing protocols used in the EU to discriminate classical scrapie from other TSEs, notably atypical scrapie and BSE, is shown in Figure 5.3. In the United States, the official test currently used for scrapie diagnosis is IHC. Western blot and ELISA may be used as supplemental tests or when tissues are not suitable for IHC.
The presence of the scrapie agent in lymphoreticular tissue in sheep and goats offers the opportunity for the diagnosis of scrapie in live animals using IHC, because some lymphoreticular tissues, such as those found in tonsil, third eyelid (nictitating membrane), and rectal mucosa, can be readily obtained from the live animal by non-invasive means. O'Rourke et al. (2000) described the technique for antemortem biopsy of nictitating membrane by using topical anesthetic and a restraint apparatus. Schreuder et al. (1998) have described the procedure for tonsillar biopsy in anesthetized sheep for collection of palatine tonsil for pre- clinical diagnosis of scrapie. PrPSc may be identified in biopsies of rectal mucosa (Espenes et al. 2006) and can be readily obtained just 1 cm inside the anus using a local anesthetic of 2% xylocaine gel and a speculum, with the ventral floor cut away to access the rectal mucosa (Raksa et al. 2016). Through use of tonsillar biopsy, preclinically affected animals were identified one and a half years before clinical signs normally appear in sheep of known genotypes with predictable incubation periods.
Figure 5.3 Testing flowchart for transmissible spongiform encephalopathies (TSEs) in small ruminants. BSE, bovine spongiform encephalopathy; ELISA, enzyme - linked immunosorbent assay. Source: Adapted from TSE Strain Characterisation in Small Ruminants: A Technical Handbook,Version 2.0, January 2019. London: Animal and Plant Health Agency.
While this represents a significant breakthrough in the antemortem diagnosis of scrapie, techniques that depend on identification of the scrapie agent in lymphoreticular tissue must be used with caution, as there is a significant potential for false-negative results. This is because the presence of PrPSc in lymphoreticular tissues of infected animals depends on a number of factors, including the age of the animal, its susceptibility to the disease based on genotype, the strain of the scrapie agent involved, and possibly other factors yet unknown that may impact the distribution of the scrapie agent in tissues. In the study of Schreuder et al. (1998), for example, there was a difference in the presence of PrPSc in animals between 3.5 and 4 months of age. Similarly, Monleon et al. (2005) compared the presence of PrPsc in CNS tissue and lymphoreticular tissue at post mortem in sheep of a specific genotype in a known infected herd, and reported that in some preclinical cases the agent was present in tonsil and lymph node but not in brain, while in some advanced and terminal clinical cases the reverse was true, with the agent present in brain but not in tonsil or lymph node. Monleon et al. (2011) later conducted similar studies on rectal mucosa and third eyelid biopsies in both sheep and goats, and found that the sensitivity of rectal mucosa and third eyelid assays was 36% and 40%, respectively, relative to infection status established at post mortem. A low number of lymphoid follicles present in the biopsy samples may have contributed to the occurrence of false-negative results. For regulatory purposes in the United States, rectal biopsies are preferable to third eyelid biopsies, because they typically have more lymphoid follicles and therefore result in fewer invalid tests (USDA 2020). Also, as a practical matter, it is easier to collect rectal biopsies than third eyelid biopsies and the test can be easily repeated later if necessary.
Currently, the most reliable diagnosis of animal TSEs is based on postmortem examination of brain and LRS tissues (Konold et al. 2020). In the EU, scrapie and BSE surveillance projects are based on examination of brain tissues (Commission regulation [EC] No. 999/2001). However, new assays are being developed that can amplify PrPSc prions in vitro in a principle analogous to PCR. Currently two such assays are available: prion misfolding cyclic amplification (PMCA) and real-time quaking induced conversion (RT-QuIC) (Saborio et al. 2001; Atarashi et al. 2008). Both methods are more sensitive compared to currently available diagnostic tests. These techniques could be used in future for antemortem diagnosis provided that they are validated, taking in consideration appropriate sampling procedures and knowledge of disease pathogenesis in specified hosts.
At necropsy, gross findings, if present at all, are limited to evidence of cachexia and possibly hair loss and skin lesions, if pruritus is a component of the clinical picture. However, postmortem histologic examination of the CNS remains an important element in the definitive diagnosis of scrapie. The principal lesion is a progressive degeneration of the gray matter, found most commonly in the thalamus, midbrain, cerebellar cortex, and medulla oblongata, but uncommonly in the cerebral cortex and spinal cord (Hadlow 1961). The most consistent site for finding morphologic vacuolar changes indicative of classical scrapie is the medulla oblongata, in samples taken at the level of the obex (Wood et al. 1997). This is not the case for atypical scrapie, where lesions are more overt and consistent in the cerebellum, thalamus, substantia nigra, and basal ganglia (Moore et al. 2008). Therefore, the medulla and cerebellum should both be examined as a minimum for thorough diagnosis and classification.
For clinically suspect cases, it is recommended to remove the entire brain for testing, while for the purpose of active surveillance, collection of samples of both the medulla oblongata and the cerebellum through the foramen magnum can be performed using a specially designed sampling spoon. Protocols for proper sample taking, handling, and submission are described in detail elsewhere (APHA 2019a).
Vacuolation in the neuropil or in neurons is considered the most significant finding. In healthy goats, vacuolation in histologic sections of the medulla is observed rarely, while vacuoles are seen in large numbers in the medulla of goats with advanced scrapie (Pattison et al. 1959). There is a notable absence of inflammatory response. The spongiform appearance of the brain classically described in sheep is less commonly observed in goats (Hadlow et al. 1980).
The absence of lesions is clearly not evidence of the absence of scrapie infection, because infection can exist in the absence of either clinical signs or pathologic changes.
Detectable PrPsc precedes vacuolation and clinical signs, making the immuno-based tests a more sensitive option. For this reason, IHC on tissue sections and/or immunob- lotting/ELISA methods on fresh tissues to demonstrate accumulation of PrPSc should be carried out in parallel with routine histology in suspected cases. This is also recommended where lesions are mild in severity and considered equivocal.
Diagnosis
Clinical diagnosis of scrapie is presumptive on the basis of slowly progressive neurologic disease in an adult goat, and is confirmed by identification of characteristic lesions histologically in the CNS and the presence of PrPSc by IHC or immunoblotting in nervous or lymphoreticular tissue. In some cases, mouse inoculation studies may be carried out using tissue from suspect cases to confirm the diagnosis and differentiate classical scrapie from atypical scrapie or BSE, if this distinction cannot be achieved unequivocally using current in vitro diagnostic methods, mainly western blot and IHC.
If chronic pruritus is a component of the clinical picture in goats, differential diagnoses should include lice, mange, paraelaphostrongylosis, elaphostrongylosis, and primary or hepatic photosensitization. If the onset of pruritus is rapid and the course of disease short, then pseudorabies and rabies should be considered. When tremors and progressive incoordination are the predominant signs, CAE, pregnancy toxemia, enzootic ataxia, parelaphostrongylosis, elaphos- trongylosis, gid, spinal cord abscesses, Borna disease, and the dumb form of rabies should be ruled out. However, none of these conditions is likely to be as slowly progressive as scrapie. If circling is present, listeriosis should be ruled out. Unexplained weight loss with no other localizing signs can be consistent with scrapie in goats. The differential diagnosis of unexplained weight loss is discussed in Chapter 15.
Treatment
There is no treatment for scrapie. Even if therapeutic options were available, there would likely be little or no opportunity to use them, because scrapie has become a reportable and highly regulated disease worldwide. Suspect cases are increasingly likely to be euthanized for diagnostic purposes by the appropriate authorities. Nonetheless, concerns about TSEs in human beings have prompted research into possible therapeutic regimens that in principle could be applicable to animals with scrapie. Obstacles and opportunities for treatment of TSEs have been reviewed (Liberski 2004; Weissmann and Aguzzi 2005). Given the potential public health implications, the policy emphasis is to control and, if possible, eradicate animal prion diseases rather than treat them.
Control
Control of scrapie at the herd level involves improved awareness of the disease among producers and implementation of appropriate management practices to minimize the risk of infection. Goat owners need to be educated about the insidious nature of scrapie, to be able to recognize the potentially varied clinical signs of disease, and to understand their regulatory responsibilities around existing control programs. In terms of improved management, the risk of scrapie can be reduced in herds by minimizing outside purchases of stock and reducing opportunities for horizontal transmission. If outside purchases are considered, then genotyping should be carried out as a condition of purchase with only resistant genotypes allowed to enter the herd. Sheep and goats should not be commingled when the infection status of sheep is unknown. During kidding season, placentas and other fetal membranes should be found and removed immediately for burial or incineration. Bedding should be changed in kidding pens between kid- dings and the soiled bedding also buried or incinerated. Any animals showing tremors, incoordination, or pruritus should be isolated from the herd and a definitive diagnosis attempted. There are currently no vaccines available for controlling scrapie.
Historically, selection for genetic resistance to scrapie was a control tool available only to sheep producers, but knowledge of the relationship of genotype to resistance in goats has advanced sufficiently that the option has become available to goat owners as well. As discussed earlier, three alleles of the prion protein gene, K222, D146, and S146, are now known to confer genetic resistance to classical scrapie in goats. In the United States, some labs are now offering genetic testing for goats to assist with selective breeding programs around scrapie control. The University of California at Davis (UC Davis 2020) will perform the necessary DNA tests on a sample of 20-30 hairs with roots intact and reports results within 6-10 days, using the interpretation as shown in Table 5.5.
Goat owners considering genetic selection as a control option need to keep some things in mind. Studies in the United States have demonstrated that both S146 and K222 may occur in various goat breeds, but at notably different frequencies. The S146 allele was identified in 7 out of 10 breeds of both meat and dairy types, including Boer, Tennessee fainting goats (myotonic), Nubian, Alpine, Saanen, LaMancha, and Pygmy goats. The frequency ranged from 35.2% in Boer goats to 1.4% in Alpine goats and was most common in the Boer and Nubian breeds. The K222 allele was only identified in two of the dairy breeds, Toggenburg and LaMancha, and at low frequencies of 5.4% and 1.0%, respectively (White et al. 2008). Similar results were found when 10 goat breeds were evaluated in Great
Table 5.5 Interpretation of DNA testing results for resistance to classical scrapie in goats in relation to the 222 and 146 alleles ofthe prion gene.
Allele 146 Allele 222 Interpretation of results
| NN | No copies of protective variants. Susceptible to classical scrapie. | |
| NN | QK | One copy of K222 protective variant. Increased resistance to classical scrapie. |
| NN | KK | Two copies of K222 protective variant. Increased resistance to classical scrapie. |
| NS | One copy of S146 protective variant. Increased resistance to classical scrapie. | |
| NS | QK | One copy each of S146 and K222 protective variants. Increased resistance to classical scrapie. |
| SS | Two copies of S146 protective variant. |
Increased resistance to classical scrapie.
K, lysine; N, asparagine; Q, glutamine; S, serine; where serine at 146 confers resistance to scrapie and lysine at 222 confers resistance to scrapie.
Source: UC Davis 2020 / The Regents of the University of California.
Britain. The S146 PrP allele was mostly limited to the Boer breed and found at a high frequency. In contrast, the K222 allele was found in all the major dairy breeds, but at a very low overall frequency of 0.6% (Goldmann et al. 2016).
The low frequency of protective haplotypes, particularly in dairy breeds, means that selection pressure will be high and selective breeding could have an adverse effect on genetic diversity. Currently it is not known what effect selection for scrapie resistance might have on other desirable genetic traits such as milk production. Therefore, the motivations and economic advantages for voluntary selective breeding for scrapie resistance at the herd level must be clear to breeders and producers and the efforts carefully monitored for unintended effects. It is likely that in the future semen will be available from bucks with scrapieresistant haplotypes for artificial insemination, so that owners will not have to depend solely on their own bucks for developing genetic resistance in their herds. At the national level, once scrapie becomes enzootic in a country, it is difficult to eliminate. However, as the experience of Australia and New Zealand in the 1950s and South Africa in the 1960s demonstrated, if scrapie is introduced with imported stock, early recognition and aggressive stamping out measures can effectively eliminate the disease before it becomes established and strict border controls and importation requirements can keep it out. The scrapie status of many sheep- and goat-producing countries in the world remains unknown at present, due to the absence of passive and active surveillance programs.
Where the disease is enzootic, scrapie control at the national level depends on the establishment and implementation of strong regulatory programs that are acceptable to and supported by sheep and goat producers. In the United States, several approaches to sanitary control have been implemented since 1952. They have been modified in response to changing views on, and understanding of, the factors involved in scrapie transmission. Historically, these approaches have included depopulation of infected flocks, source flocks, and exposed flocks; slaughter of infected animals and all their blood relations regardless of flock of origin; slaughter of female blood relations and lateral contact sheep; and slaughter of female blood relations only. None of these approaches was completely successful.
In October 1992, as a result of a negotiated rule-making process between government, industry, and producer groups in the United States, a new Voluntary Scrapie Flock Certification Program was enacted as part of the National Scrapie Eradication Program (NSEP). This voluntary program allowed flocks to be certified as scrapie free after a five-year period. Voluntary participation involves careful animal identification and record keeping, constraints on animal movement and purchase, and regular monitoring of flocks by Animal and Plant Health Inspection Service (APHIS) veterinarians. The program applies to goats and sheep.
In 2001, APHIS announced the accelerated NSEP, with the stated goal of eradicating scrapie in the nation's sheep and goat flocks by 2010 and having official OIE recognition of the United States as being scrapie free by 2017. The key elements of the accelerated NSEP included identifying pre- clinical infected sheep through live animal testing and active slaughter surveillance; making it possible to effectively trace infected animals to their flock/herd of origin through new identification requirements; and providing effective cleanup strategies that allow producers to stay in business, preserve breeding stock, and remain economically viable.
To facilitate effective cleanup strategies, USDA/APHIS provides indemnity for high-risk, suspect, and scrapiepositive sheep and goats that owners agree to destroy; scrapie live animal testing; genetic testing in sheep; and testing of exposed animals that have been sold out of infected and source flocks/herds.
The NSEP regulations were updated in 2019 (USDA 2019). The new regulations formalize the policy of using genetic testing to identify genetically resistant or less susceptible sheep for exemption from destruction and as qualifying for interstate movement. Genetic testing in goats, however, has not yet been integrated into the NSEP. In addition, the new regulations make the identification and recordkeeping requirements for goat owners consistent with those already required for sheep owners. Producers of goats for meat or fiber and slaughter goats more than 18 months of age are now required to officially identify their animals to their flocks of birth or flocks of origin, and to maintain certain identification records for five years. The expanded requirement for goat identification is intended to allow a greater level of surveillance for goats than was previously achievable, with the goal of achieving national scrapie-free official status with the OIE. More detailed information about the NSEP is available online at https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/ animal-disease-information/sheep-and-goat-health/ national-scrapie-eradication-program.
In the EU, scrapie has been a notifiable disease since January 1993. Active surveillance programs for scrapie in sheep and goats were legislated in the EU in 1998, and in 2001 regulation (EC) No. 999/2001 of the European Parliament established rules for the prevention, control, and eradication of certain TSEs in the European community, including scrapie. Genotyping of confirmed positive sheep was mandated in this legislation and rules governing the import and export of sheep and goats and their products were spelled out.
Until 2003, it was left to member states to determine what to do with herds and flocks from which scrapie had been confirmed. However, Commission Regulation (EC) No. 260/2003 set community-wide rules for the compulsory disposition of known positive flocks and herds. All animals in goat flocks were to be destroyed. In sheep flocks, there was an option to genetically test and cull susceptible genotypes only. However, in June of 2020, based on the accumulated evidence that K222, D146, and S146 alleles confer genetic resistance against classical scrapie strains known to occur naturally in the EU goat population, a new EU regulation, 2020/772, was adopted, amending various annexes to the original TSE regulation (EC) No. 999/2001 with regard to scrapie eradication measures in goats. Under EU regulation 2020/772, if a goat is confirmed with classical scrapie, only those goats in the herd that are genetically susceptible to scrapie need be destroyed, thus aligning the approach to that already existing for sheep.
Enhanced surveillance activities associated with programs for the eradication of classical scrapie have resulted in the increased detection of cases of atypical scrapie. However, from the regulatory standpoint, atypical scrapie is considered a separate and distinct disease that does not trigger the regulatory interventions associated with detection of classical scrapie. Consistent with the OIE position, countries generally do not restrict or depopulate animals exposed to atypical scrapie, as it is considered to be clinically, pathologically, biochemically, and epidemiologically unrelated to classical scrapie. It is not considered to be contagious and more likely represents a spontaneous degenerative condition of older sheep and goats.
Bovine Spongiform Encephalopathy
BSE, like scrapie in goats and sheep, is one of the prion- associated diseases classified as a TSE. BSE was first recognized as a disease of cattle in the United Kingdom in 1986 (Wells et al. 1987). Then, in 1996, it was reported that a new, fatal, human TSE, called vCJD, was thought to be associated with the consumption of contaminated animal products derived from BSE-infected cattle (Bruce et al. 1997; Collinge et al. 1996). Thus, BSE became a zoonotic disease, a major public health concern, a catastrophe for the cattle industry, and the focus of enormous international media attention.
There are two reasons for discussing BSE in this textbook of caprine medicine. First, because BSE can be produced in goats by experimental inoculation, and second, because in 2005, a naturally occurring case of BSE was identified through active surveillance in a goat from a commercial dairy herd in France (Eloit et al. 2005). This triggered retrospective studies in several EU countries and as a result another goat, which was originally diagnosed as scrapie in Scotland in 1990, was subsequently confirmed as BSE (Jeffrey et al. 2006; Spiropoulos et al. 2011). In the experimental infections and the single natural case from Scotland that showed signs of disease, the clinical presentation was indistinguishable from scrapie (Foster et al. 2001; Jeffrey et al. 2006).
As a result of the naturally occurring cases, surveillance activities of small ruminants in Europe were sharply increased, but as of 2020 no additional cases of BSE in goats have been reported from Europe or elsewhere in the world. Nevertheless, since there are still unresolved questions concerning the origins of BSE and its epidemiology, practitioners should be aware that goats are susceptible to BSE and that it cannot be differentiated from scrapie based on clinical signs, nor diagnosed definitively in the live animal.
A comprehensive discussion of BSE, which remains primarily a bovine disease, is beyond the scope of this book. The information presented here is limited to what is relevant and known about the disease in goats and its implications for the goat industry. A more detailed discussion of BSE and its diagnosis and control in cattle can be found elsewhere (OIE 2018b, Simmons et al. 2018).
Etiology and Pathogenesis
Like scrapie, BSE is a TSE and the generally accepted cause of BSE is the expression of an abnormal form of a naturally occurring, cell membrane-associated, prion protein called PrPC, which is a ubiquitous protein found in cells throughout the host animal. It is believed that the abnormal conformation of this normal cellular protein is triggered by the introduction into the host animal of an infectious form of the PrP protein, known as PrPSc (PrP- scrapie), but also referred to in the literature as PrPres (protease resistant) or PrPd (disease associated). It is proposed that during the infectious process this abnormal isoform of the prion protein binds to and changes the normal PrPC three-dimensional conformation, with at least a portion of the PrPc-associated α-helical structure assuming the form of β-sheets that is characteristic of PrPSc. In many cases the accumulation of this abnormal PrPSc in the gray matter of the brain leads to a histologically apparent degeneration characterized by sponge-like vacuolation of affected brain tissue that gives the disease its descriptive name. Alternate theories on the pathogenesis of TSEs also persist, for example that the wall-less bacterium Spiroplasma may be a transmissible agent of TSEs (Bastian et al. 2007, 2011).
It is well documented in sheep that the genotype of the host animal plays some role in the animal's susceptibility or apparent resistance to scrapie infection. Results of experimental infection of sheep with BSE indicate that these genetic factors may also influence the ovine response to BSE, though a recent study demonstrated that sheep with the most scrapie-resistant genotype ARR/ARR could be orally infected with BSE and have significant amounts of PrPSc in their spleens (Andreoletti et al. 2006). In goats, a recent European review of both field and experimental data considered the evidence robust enough to conclude that the K222, D146, and S146 alleles confer genetic resistance against classical scrapie strains known to occur naturally in the EU goat population, but that there was insufficient evidence to assess the impact of these alleles on susceptibility to atypical scrapie and BSE (Ricci et al. 2017).
Epidemiology
BSE was first recognized in cattle in the United Kingdom in 1986. It manifested as a neurologic disease with characteristic behavioral changes that led to the popular name of “mad cow” disease. While the cause of the disease was unknown, epidemiologic studies strongly suggested that the disease was being spread through the feeding of concentrate feed that contained animal protein byproducts derived from ruminants. It was hypothesized that removing ruminant protein from cattle feed supplements would lead to a reduction of disease occurrence, and in fact this turned out to be correct (Wilesmith et al. 1992; Anderson et al. 1996). A ban on feeding ruminant-derived meat and bone meal (MBM) to cattle in the United Kingdom was instituted in 1988. Following that ban, the UK epizootic peaked in 1992 with 37 280 confirmed cases, and has declined steadily since, with only 225 cases confirmed in 2005 and no cases in 2016. There have been five cases of confirmed BSE in the United Kingdom since 2014, with the most recent one being identified in September 2021. All of these five cases were in fallen stock not destined for the human food chain (Government of the United Kingdom 2021).
No other food-producing animals were recognized as manifesting signs of BSE during the course of the UK BSE epizootic, though retrospectively one goat in Scotland originally diagnosed with scrapie in 1990 was confirmed as a case of BSE (Spiropoulos et al. 2011). In zoologic parks, some exotic ruminants that likely received prion- contaminated livestock feed became infected with a TSE. These included greater kudu, nyala, eland, gemsbok, and oryx. Similarly, 20 captive exotic felids, including cheetahs, lions, ocelots, pumas, and tigers, developed TSEs, most likely from being fed meat or offal from BSE-infected cattle. In addition, 89 domestic cats diagnosed with feline spongiform encephalopathy (FSE) in the United Kingdom were associated with the BSE epizootic between 1989 and 2001 (APHA 2019b). Several other countries have also detected cases of FSE, including Norway, Liechtenstein, France, Portugal, Italy, and Switzerland.
Unfortunately, livestock feeds containing MBM had also been exported to other countries, as had infected livestock from the United Kingdom, and ultimately additional cases of BSE were found in numerous countries in Europe and elsewhere, though never to the extent that occurred in the United Kingdom. By the end of 2006, 24 countries had reported at least one case of BSE. This included 20 countries in Europe as well as Israel, Japan, Canada, and the United States. As a result, the EU member states and many other nations instituted bans on feeding ruminant-derived protein to ruminant livestock. In Europe, problems with cross-contamination of feed for ruminants and nonruminants prompted the bans to include prohibiting the feeding of most animal proteins to food-producing animals. As experimental studies and field observations indicate that horizontal transmission does not occur from cow to cow, the feed ban is the most effective tool there is to control the spread of the disease in livestock. Studies have also been done to determine whether BSE can be transmitted by frozen cattle semen and frozen cattle embryos. This does not appear to be a risk. Similarly, researchers were unable to transmit BSE via goat embryos (Foster et al. 1999).
The first human death due to vCJD occurred in a 19-year-old British man in 1995 and by 1996 the likely link between BSE and vCJD was acknowledged. It became accepted that consumption of bovine meat products that were contaminated with nervous tissue of infected cattle was the most likely route of transmission of BSE prions to humans in cases of vCJD. As a result, the EU member states and other countries with BSE-infected cattle instituted new restrictions on the bovine slaughter industry to exclude so-called specified risk materials (SRM) from meat products derived from slaughter plants.
The SRM are tissues with the potential to contain significant amounts of BSE infectivity in infected animals. This SRM restriction was not instituted in the United States until 2004, since the first bovine case of BSE in the country was only diagnosed in 2003. This intervention appears to have had the desired effect of limiting the occurrence of vCJD. The annual incidence of vCJD in the United Kingdom peaked in 2000 at 28 cases and has declined since. From 1995 through October 2019, a total of 232 cases of vCJD were reported worldwide, with only 7 of those cases occurring since 2012 (Creutzfeldt-Jakob Disease International Surveillance Network 2019). A total of four cases have been confirmed in the United States, the most recent in 2014.
The origin and character of the infective prion initially consumed by cattle to produce the epizootic of BSE in the United Kingdom have never been established. One of the original theories is that the infective prion was originally the scrapie prion, introduced into the food chain via infected sheep or goat tissues, and that the prion was somehow transformed into a unique species-specific agent affecting cattle. An alternate theory is that a spontaneous mutation of normal bovine prion with the capacity for infectivity entered the animal food chain via affected slaughter cattle from which MBM was derived. This theory is now being examined in greater depth since the finding of atypical manifestations of BSE in cattle (Biacabe et al. 2004; Casalone et al. 2004; Brown et al. 2006) and the reported capacity of one atypical BSE prion strain to undergo transformation after serial passage in mice, to produce a neuro- pathologic and molecular disease phenotype that was indistinguishable from that of mice infected with the classical BSE prion strain (Capobianco et al. 2007). Regardless of the origin, the prion that produces classical BSE can be differentiated from the two forms of atypical BSE, H-type BSE and L-type BSE, as well as from various strains of the scrapie prion, as a result of improved laboratory techniques available for characterization of prion proteins in tissues from necropsy or slaughter.
The clinicopathologic similarities of BSE and scrapie spurred research on prion diseases in small ruminants, including investigations on the potential of sheep and goats to become infected with BSE. Successful experimental infections demonstrated that both sheep and goats are susceptible to BSE infection and can manifest clinical disease. The first experimental transmission of BSE to goats was reported by Foster et al. (1993). Six goats were inoculated with brain homogenates of known BSE-infected cows either intracerebrally or orally. Three of three Anglo- Nubian goats challenged intracerebrally developed clinical signs of BSE 506-570 days after challenge and two of three goats challenged orally developed the disease 941 and 1501 days after challenge (Foster et al. 1993, 2001). This indicated that the presumed natural route of infection, i.e., oral ingestion, could produce BSE in goats.
The results of such studies, and the general concern for the zoonotic potential of TSEs, prompted the establishment and/or expansion of active surveillance programs for the presence of BSE and scrapie in cattle, sheep, and goats. In May 2001, the European Parliament approved EC Regulation No. 999/2001 (European Commission 2001), laying down rules for the prevention, control, and eradication of certain TSEs. This comprehensive legislation addressed numerous key issues: establishment of a national status category for countries relative to the presence of TSE based on quantitative risk analysis; establishment of active surveillance programs for TSEs in sheep and goats (scrapie and BSE) with minimum sample sizes identified for each member state; identification of SRM for small ruminants and cattle; establishment of regulatory procedures for the diagnosis of TSE and destruction of sheep and goat flocks confirmed with either scrapie or BSE, along with guidelines for compensation of owners; rules for import and export of live animals, semen, embryos, and animal products relative to the TSE status of importing and exporting countries; identification of TSE reference laboratories; and identification of approved sampling and laboratory methods.
Beginning in 2002, the EU countries started to test sheep and goats in large numbers for TSEs. The samples included healthy animals sent to slaughter, animals culled as part of ongoing scrapie eradication programs, reported suspect TSE cases, and other suspect animals such as fallen stock or animals that had died on farm. With regard to goats, the EU countries tested 54 444 in 2002, 63 022 in 2003, 36 115 in 2004, and 265 489 in 2005 (European Commission 2003, 2004, 2005, 2006a).
That increased surveillance produced a notable result. On January 28, 2005, the European Commission announced that an apparently healthy 2.5-year-old goat slaughtered in France in October 2002 and tested under the active TSE surveillance system had been identified as a positive BSE case. Mouse bioassays confirmed that this represented the first known case of naturally occurring BSE in a goat (Eloit et al. 2005). As a precaution, the French authorities culled the entire herd of origin; no animals or animal products were allowed to enter the human or animal food chains. All the other approximately 300 adult goats in the herd were tested, with no detectable evidence of a TSE found.
In February 2005, UK authorities announced a possible second case of naturally occurring BSE in a goat (Eurosurveillance 2005). This dairy goat, from Scotland, was born in 1987 and may have been fed a BSE- contaminated calf weaner mix. It was examined for a complaint of neurologic signs and pruritus in 1990 and diagnosed with scrapie at that time. Samples from the goat had been selected for reevaluation as part of a research effort to assess newer, more sensitive testing methods for discriminating scrapie from BSE in goats, and BSE was tentatively diagnosed based on that reexamination (Jeffrey et al. 2006). The diagnosis of BSE was subsequently confirmed by mouse bioassay (Spiropoulos et al. 2011).
These two confirmed cases established that naturally occurring BSE can occur in the goat. However, unanswered questions remained about the origin of these infections, the overall susceptibility of goat populations to BSE, and the risk that goats and goat products may act as a source of infection for vCJD in humans.
The confirmed BSE-positive French goat was born before the EU initiated its extensive animal protein feed ban in 2001, so the possibility exists that oral consumption of the BSE prion in contaminated feed was the source of exposure for this goat. Similarly, the UK goat was born in 1987, before the 1988 UK feed ban on MBM, and is believed to have consumed BSE-contaminated feed. Even before the emergence of these two reports, there was considerable concern among experts that BSE would be identified in small ruminants, though the focus of concern was more on sheep than on goats, due to the greater numbers of sheep in Europe and the higher economic value of the sheep industry (Schreuder and Somerville 2003).
The Scientific Steering Committee (SSC) of the European Commission issued an opinion on the risk of infection of sheep and goats with the BSE agent in 1998 (European Commission 1998). In summary, the committee believed that the transmission of the BSE agent to small ruminants was possible and, if it occurred, it would likely be through the same mechanism by which cattle were infected, namely by consumption of concentrate feed containing prion- contaminated animal protein. Because the route was likely to be the same, the geographic distribution of small ruminant cases would also likely be the same, with the occurrence in small ruminants reflecting the occurrence in cattle. It was also recognized that dairy goats were more likely to have consumed contaminated concentrate feed, with a possible higher risk of exposure to prion contamination than sheep and goats kept for other purposes. By 1998, it was already clear that the feed ban on animal protein in the United Kingdom had effectively dampened the BSE epizootic in cattle there, with fewer cases being reported each year. While it was reasonable to assume that the risk of infection in small ruminants would subside in a similar fashion, the SSC expressed concern that it was not known if BSE in small ruminants would behave like scrapie in sheep, or like BSE in cattle.
In BSE of cattle, there is no indication that the disease is transmitted either horizontally or vertically between animals. Virtually all bovine cases are believed to derive from direct consumption of BSE prion-contaminated feed. In contrast, scrapie can be transmitted between small ruminants, either directly or indirectly. Thus, if BSE were to behave like scrapie in small ruminants, the infection could continue to be maintained in sheep and goat populations and continue to spread despite the feed ban.
One experimental study has shown that BSE is able to spread from infected ewes to lambs, though it was not clear if this occurred in utero or in the perinatal period (Bellworthy et al. 2005a). However, the fact that only two goats and no sheep have ever been confirmed with naturally occurring BSE since the SSC issued its opinion in 1998 suggests that BSE has not been widely circulated or maintained in European small ruminant populations, which were at high risk.
The other major concern is the possible risk to humans of BSE occurring in small ruminants. The issue is whether or not the controls now in place to protect humans from zoonotic prion infection from bovine products are sufficient to protect humans from BSE in small ruminants. Again, this would depend on whether or not BSE in small ruminants behaves like scrapie in small ruminants or like BSE in cattle. The distribution of prions in host tissues is the key factor for consideration in this regard. Several studies have now been completed on the experimental infection of sheep with BSE (Foster et al. 1993, 2001; Jeffrey et al. 2001; Bellworthy et al. 2005b). These studies indicate that experimental BSE in sheep behaves like scrapie, with a wide distribution of PrPsc in tissues of the LRS and the enteric nervous system in the preclinical stages of infection. Among the PrPsc-positive tissues identified were submandibular, retropharyngeal, prescapular, mesenteric, ileocecal, and mediastinal lymph nodes; tonsil; and spleen. In the enteric system, PrPsc was identified in association with lymphoid tissue or enteric nervous tissue in the large and small intestine. Of interest was the observation of PrPsc in the abomasum (Jeffrey et al. 2001). This PrPsc was found in macrophages associated with focal inflammatory lesions, presumed to be induced by gastrointestinal parasites.
The notion that PrPsc-laden macrophages can be drawn to sites of infection as part of the inflammatory response suggests the possibility that in cases of mastitis, even sub- clinical mastitis, macrophages containing PrPsc could be drawn to the udder and thereby be present in milk. In fact, this process has been observed in mastitic sheep with scrapie (Ligios et al. 2005). Scrapie infectivity in milk derived from sheep and goats affected by scrapie can be detected in the absence of mastitis (Konold et al. 2008, 2016). However, taking into consideration the low infectivity levels detected in the milk and the interspecies transmission barrier, it has been suggested that the risk associated with consumption of ewe or goat milk by humans certainly remains limited (Lacroux 2008). Clearly, classification of milk as an SRM would have devastating consequences for dairying in small ruminants. The fact that prions are not destroyed by standard methods of pasteurization adds to that concern.
Following the confirmation of the BSE-positive French goat, and with these concerns in mind, the EFSjA undertook risk assessments on the possible transmission of BSE through consumption of goat products. While acknowledging that the available data on BSE in small ruminants was limited, the scientific panel concluded that in light of current scientific knowledge and irrespective of their geographic origin, milk and milk derivatives (e.g., lactoferrin, lactose) from small ruminants were unlikely to present any risk of TSE contamination, provided that milk was sourced from clinically healthy animals. Exclusion of animals with mastitis was considered to reduce the potential risk, and further assurance of healthy milk could include milk tests for total somatic cell counts indicative of inflammation (EFSA 2004). With regard to goat meat and goat meat products, EFSjA scientists again cited a lack of sufficient data on infectious load, potential distribution of BSE in goat tissues, prevalence of infection, species barriers, and levels of human goat meat consumption to draw definitive conclusions. Despite that caveat, their report stated that there was no evidence of a link between goat meat consumption and a higher risk of vCJD, based on epidemiologic data from the United Kingdom indicating that the occurrence of vCJD in certain ethnic groups likely to consume goat meat at a higher level was no different than that of the general population (EFSA 2005).
Since these reports and the identification of the BSEpositive goat from France in 2005, well over 2 million goats from 28 EU member states and 4 non-EU member states in Europe were tested for TSEs between 2006 and 2018, under the EU TSE Surveillance program (EFSA 2019). Through this effort, approximately 12 000 goats were identified with classical scrapie. All these classical scrapie cases in goats were subject to discriminatory testing for BSE, and not a single additional case of caprine BSE was detected. This accumulating data on the absence of BSE in goats in Europe, which was the epicenter of BSE in cattle, is reassuring. It suggests that the ban on ruminant protein in feed, which was highly effective in controlling BSE in cattle, also had a positive impact on curtailing BSE infection in goats and sheep.
At present, the OIE, which is the standard-setting body for the World Trade Organization concerning safe trade in animals and animal products, does not consider small ruminants to be epidemiologically significant in the transmission of BSE. The OIE Terrestrial Animal Health Code states that with regard to the importation of live animals, cattle pose the only known risk, though other species remain under study (OIE 2021b).
Clinical Findings and Diagnosis
Naturally occurring BSE has been diagnosed in only two goats. One was clinically normal at slaughter; the second had shown neurologic signs and pruritus consistent with the diagnosis of scrapie made at the time of examination. Clinical BSE has been produced in goats by experimental infection and signs of the experimentally induced disease have been described (Foster et al. 1993, 2001; Konold et al. 2010). These descriptions should be understood in the context in which they were produced. The routes of infection and dosages of infective material used under experimental conditions may not produce the clinical manifestations of the disease as likely to be seen under natural circumstances, particularly with regard to the temporal course of clinical disease, which appears relatively short when compared to natural BSE in cattle or natural scrapie in goats and sheep.
Foster et al. (1993, 2001) reported that three goats inoculated intracerebrally with a brain homogenate from BSE- infected cattle developed clinical disease between 506 and 570 days post inoculation. Two of three goats dosed orally developed clinical disease at 941 and 1501 days, respectively, and a third remained healthy at 1720 days post challenge. All three intracerebrally inoculated goats and one of the orally dosed goats developed sudden and pronounced ataxia, which progressed so acutely, once manifest, that each of these goats had to be euthanized within six days after the onset of ataxia for reasons of welfare. The other orally dosed goat that developed clinical signs did not show signs of ataxia. Rather, it simply became lethargic and lost weight over a period of three weeks before it was culled. None of the five affected animals developed pruritus, a sign characteristic of scrapie, the more common TSE of small ruminants.
Konold et al. (2010) infected five goats intracerebrally with a homogenate of bovine BSE brain stem. Seven clinical signs were recorded: aversive reaction to approaching the head; repeated startle response; positive scratch test (pruritus); ataxia; abnormal posture; head or body tremor; and absent menace response. The earliest sign occurred in one goat at 15 months post inoculation, but the majority of signs occurred in the five goats between 24 and 34 months post inoculation. Only one goat showed all seven signs, but all goats showed at least four. Notably, three of the five showed signs of pruritus, previously not reported in goats experimentally infected with BSE, and three showed an absent menace response. It was recommended that signs assessed for the clinical diagnosis of BSE in goats should include postural and gait abnormalities, pruritus, and visual impairment.
Given the uncertainty of the clinical presentation of BSE in goats and given the serious regulatory concerns associated with all TSEs in ruminant animals, it is prudent for veterinary practitioners to consider both of the TSEs, scrapie and BSE, in the differential diagnosis of neurologic disease in goats, particularly chronic or progressive neurologic disease. It is reasonable to assume that were clinical BSE to occur naturally in goats, it would appear similar to BSE in cattle or scrapie in goats and sheep.
In cattle with natural BSE, the onset of disease is insidious and the course is long. The predominant clinical signs are neurologic in nature and include apprehension, hyperesthesia, and ataxia, the latter progressing to the point at which affected animals may be unable to rise. It is important to note that clinical signs of the TSEs may be very subtle and often go unnoticed until near the end stage. The clinical disease in cattle is described in more detail elsewhere (Constable et al. 2017). The clinical signs of scrapie in goats are described in this chapter in the discussion of scrapie, along with the likely differential diagnoses for consideration in both scrapie and BSE.
Clinical Pathology and Necropsy
Currently, there are no serologic or other antemortem laboratory tests available for the diagnosis of BSE and the diagnosis must be confirmed at necropsy. There are no consistent hematologic, biochemical, or gross pathologic changes in animals with clinical or preclinical BSE.
BSE produces characteristic spongiform lesions in selected regions of the brain, similar to those described for scrapie (Foster et al. 2001). However, in small ruminants, discriminatory tests such as IHC and western blot using antibodies are required to determine if the characteristic lesions are due to scrapie or BSE. In those clinical cases where practitioners suspect that TSEs may be involved, regulatory authorities must be notified so that suitable samples of brain stem and cerebellar tissues can be collected for analysis if deemed necessary. Descriptions of the necropsy techniques for collecting appropriate tissue samples for diagnosis of TSEs in goats and sheep are available (USDA 2007c; APHA 2019a).
A three-tiered testing program is currently in effect in the EU to identify BSE in small ruminants, as illustrated in Figure 5.3. Brain stem tissue from sheep and goats is first screened in active surveillance programs for TSE using approved rapid screening tests based on the ELISA format that can detect PrPSc in samples, but cannot distinguish between BSE and scrapie. The second tier of testing involves application of discriminatory tests to TSE-positive samples, such as IHC or western blot, where new discriminatory procedures for differentiating BSE and scrapie in small ruminants have been developed. These approaches include evaluation of the molecular size and glycosylation pattern of PrPSc for western blot, and epitope mapping studies for IHC and western blot using antibodies directed at different portions of the prion molecule (Lezmi et al. 2004; Thuring et al. 2004; Jeffrey et al. 2006). Despite application of these specialized methods, it may still not be possible to exclude BSE. In this case tissues from the suspect animal are subjected to the third phase of confirmatory testing, which involves inoculation of inbred or transgenic mice and comparison of the resulting disease phenotype with that caused by BSE. Mouse inoculation studies are laborious and take upward of a year or more for the mice to develop disease and for final tests to be completed. Such delays can hamper prompt, effective regulatory action.
Treatment and Control
There is no treatment for BSE. Even if therapeutic options were available, there would likely be little or no opportunity to use them, because the disease is reportable and highly regulated worldwide. Suspect cases would be euthanized for diagnostic purposes by the appropriate authorities.
Different countries have different specific regulations for the control of BSE, but the basic principles apply in all cases. The key element in controlling the spread of BSE has been the ban on feeding ruminant-derived MBM to ruminant animals, which was later expanded in some countries to a ban on the use of most animal-derived protein in the rations of all farm animal species. The global prevalence of BSE in cattle has progressively decreased since these measures were adopted, first in the United Kingdom in 1988 and then elsewhere. In addition, countries without BSE strive to keep it out by establishing import restrictions on live ruminants and a varying array of ruminant products, including, in some cases, semen and embryos of cattle, sheep, and goats. BSE-free countries may also establish surveillance programs to ensure that the disease has not been introduced despite import regulations. The OIE sets standards for its member countries to establish an official status relative to BSE, identifying countries as having a negligible BSE risk or a controlled BSE risk. Currently 49 countries, including the United States, are officially recognized in the negligible risk category (OIE 2020).
Caprine Arthritis Encephalitis and Maedi-Visna
CAE is a disease of goats caused by a retrovirus belonging to the small ruminant Ientivirus (SRLV) serogroup. There are five main clinical presentations for CAE, namely arthritis, which is predominant; mastitis; pneumonia, a neurologic form; leukoencephalomyelitis; and progressive weight loss. The focus of the discussion here is the neurologic form of the disease. A second SRLV, known as maedi- visna virus (MVV), affects sheep and produces similar clinical conditions in that species, with the pneumonic form being predominant.
In the past, some cases of neurologic disease in goats were attributed to infection with MVV. Clinically and his- topathologically, these cases were strongly suggestive of CAE, but occurred at a time when relatively little was known about the caprine arthritis encephalitis virus (CAEV) and its relationship to MVV. The neurologic disease of goats caused by SRLVs is described below and new relevant information about the relationship of CAEV and MVV is presented. The arthritic, pneumonic, and mastitic forms of CAE are discussed further in Chapters 4, 9, and 14, respectively.
Etiology
CAEV and MVV are positive, single-stranded RNA enveloped viruses in the family Retroviridae in the genus Lentivirinae. The lentiviruses are also known as slow viruses, because they cause persistent infections that slowly manifest as progressive, clinical disease. More detailed information about the physicochemical, genetic, and immunologic aspects of the SRLVs is available in the main discussion of CAE in Chapter 4.
The lentiviruses are classified into five serogroups, reflecting the hosts with which they are associated. CAEV and MVV belong to the SRLV semigroup and cause CAE in goats and MV in sheep, respectively. The other serogroups include the cattle serogroup, associated with bovine immunodeficiency disease; the equine serogroup, associated with equine infectious anemia; the feline serogroup, associated with feline immunodeficiency disease; and the primate serogroup, which contains the simian immunodeficiency virus (SIV) and the human immunodeficiency virus (HIV). The latter is responsible for acquired immunodeficiency syndrome or AIDS in people.
The close relationship of the SRLVs to HIV spurred a great deal of research on CAEV and MVV. Though neither is zoonotic, the diseases they cause, CAE and MV, have served as models for the study of AIDS. As a result of this heightened interest, the veterinary community now has a better understanding of the SRLVs, their relationship to each other, the diseases they produce, and what distinguishes them from the primate group of lentiviruses, notably HIV (Bose et al. 2015).
MVV is the prototype SRLV, first described in the 1950s, whereas CAEV was first described in the 1970s. Historically, MVV and CAEV were considered as distinct viral entities with well-defined host specificity for sheep and goats, respectively. However, ongoing advances in phylogenetic analysis and studies of cross-transmission have rendered this view archaic, and the two viruses are now considered to represent a genetic continuum within the SRLV serogroup.
Zanoni (1998) reported phylogenetic studies of SRLVs based on available sequence data for fragments of their env, pol, and gag genes and their long terminal repeats (LTR). The results indicated that at least six different clades could be differentiated, with no clear separation of SRLV strains derived from goats (CAEV) or sheep (MVV).
Later, Shah et al. (2004a) reported additional phylogenetic studies using a larger number of virus isolates and longer sequences of the gag and pol genes, and confirmed Zanoni's observation that the SRLVs do not segregate phy- logenetically according to host specificity, demonstrating that four of nine designated subtypes of SRLV contained both sheep and goat isolates. It was also noted that Swiss strains in the so-called B1 subtype differed no more from French, Brazilian, or US strains than from each other, suggesting virus dissemination through international livestock trade. Furthermore, isolates in subtypes A3 and A4 were derived from both goats and sheep. This was documented from specific herds where there was contact between the two species, providing further evidence for interspecies transmission of the SRLV (Shah et al. 2004a, b). In a separate study, phylogenetic evidence of transmission of SRLV subtype B1 from goats to sheep in mixed flocks was also reported (Pisoni et al. 2005). Additional evidence for cross-species transmission in both domestic and wild small ruminants has since emerged, as well as new classifications of the subtypes of SRLV (Leroux et al. 2010). It appears that mutations and recombination are continuous processes that extend the genetic diversity of SRLVs and lead to the emergence of new variants, some of which may escape detection by existing diagnostic methods (Minardi da Cruz et al. 2013). Growing evidence of cross-species transmission means that some approaches to the control of CAE may need to be reconsidered, particularly where sheep and goats are commingled.
Currently, the SRLVs are classified into four genotypes, A, B, C, and E (Molaee et al. 2020), with isolates originally placed in a separate group, genotype D, now reclassified as being in genotype A. Genotype A, which includes the isolates most related to MVV, is the most heterogeneous group with at least 20 subtypes. Genotype B, which includes the isolates most related to CAEV, contains three subtypes, B1- B3. Genotype C is divided into two subtypes and refers to Norwegian isolates, while genotype E comprises subtypes E1 and E2 that were isolated in Italy.
Epidemiology
MV was first described in South Africa as a chronic respiratory disease of sheep in 1915. It was later reported from the United States in 1923 and subsequently in Iceland, where it was first recognized in the respiratory form (maedi is Icelandic for dyspnea) and then later in the neurologic form (visna is Icelandic for wasting), which occurred only in flocks of sheep with the respiratory form of the disease (Clements and Zink 1996). Intensive epidemiologic, pathologic, and virologic studies in Iceland in the 1950s led to the recognition and characterization of the respiratory and neurologic forms of the disease as being due to a single, slow virus infection (Sigurdsson 1954; Sigurdsson and Palsson 1958; Sigurdsson et al. 1952, 1957, 1960, 1962). Respiratory disease is the dominant manifestation of MVV infection. In North America, the disease is known as ovine progressive pneumonia, in France as la bouhite, in South Africa as Graaff-Reinet disease, and in Holland as zwoeger- ziekte. MVV in sheep has been reviewed elsewhere (Pepin et al. 1998). MV occurs in most major commercial sheep-producing countries except Australia and New Zealand. However, those countries do have CAE in their goat populations. The status of MV in many developing countries is unknown because surveillance programs are lacking.
Sporadic occurrences of a neurologic disease in goats characterized by granulomatous encephalomyelitis were recorded in the former West Germany in 1969 and 1978 and Sweden in 1981 (Stavrou et al. 1969; Weinhold and Triemer 1978; Sundquist et al. 1981). At the time, these were considered to be putative cases of visna or a visna-like disease of goats. Each of the German reports involved a single goat herd and the Swedish report involved multiple herds in north and central Sweden. In all these cases, a neurologic disease syndrome occurred in adult, dairy breed goats, characterized clinically by progressive paresis and histologically by two key features - granulomatous encephalitis and demyelination. In the later West German report, neutralizing antibodies to sheep visna virus were identified in affected goats (Weinhold and Triemer 1978). A visna- like virus was isolated from the Swedish goats (Sundquist 1981). Notably, the virus isolated from the Swedish goats formed syncytia in tissue culture, but was not cytopathic. This is characteristic of CAEV, whereas MVV viruses usually produce syncytia and a cytopathic effect (Querat et al. 1984).
While the neurologic form of CAE (viral leukoencepha- lomyelitis) is most common in young goats between 2 and 6 months of age, clinical cases do occur beyond this limited age range, extending well into adulthood (Norman and Smith 1983). Therefore, the occurrence of these putative visna cases in adult dairy goats is not inconsistent with CAE. In addition, adult goats with the arthritic form of the disease commonly show histologic evidence of neurologic involvement at necropsy. While CAE virus rarely induces neutralizing antibody, such antibody may develop over time (Ellis et al. 1987).
Given the better understanding of the phylogenetic relationship of some MVV and CAEV isolates and the growing evidence for cross-species transmission, it is most likely that these early cases of putative visna in adult dairy goats in Europe were actually CAE (Andresdottir et al. 2005).
New evidence on cross-species transmission of SRLV also has implications for control programs. Switzerland has a national CAE control program based on testing, culling, and indemnification that has been very effective in reducing the infection in goats from more than 50% of the Swiss goat population in the 1980s to about 0.3% in 2003 (Perler 2004). However, it has been observed that some herds participating in the program, which had been seronegative for extended periods, inexplicably became positive again. Recent investigations indicate that these herds often contain sheep or have a history of contact with sheep. Shah et al. (2004a, b) specifically demonstrated that virus obtained from goats in the control program that were commingled with sheep was phylogenetically related to the virus obtained from sheep. This means that for CAE control to be fully effective, strict segregation of goats and sheep must be a core element of the program.
Pathogenesis
While knowledge has increased considerably, some aspects of the pathogenesis are still unknown. Additional information on the transmission and early stages of infection with CAEV is provided in Chapter 4 in the section on CAE. The pathogenesis of neurologic disease caused by SRLV in goats and sheep has been reviewed (Andresdottir et al. 2005) and is summarized as follows.
Cells of the monocyte lineage in the bone marrow and spleen are the primary targets of CAEV. Replication in these cells is restricted until the monocytes differentiate into macrophages, which appears to be regulated at the transcriptional level. The mechanism by which infected macrophages cross the blood-brain barrier and enter the CNS is not fully understood. It most likely occurs via the bloodstream, with monocytes crossing non-specifically in their capacity as immune surveillance cells, or it may be in response to signals from activated T cells in response to inflammation or other infections. Other possibilities for introduction include Ientivirus infection of CNS vascular endothelial cells or choroid plexus with subsequent release of virus to the brain. Once in the brain, virus may infect a variety of cell types, and proviral DNA has been located in macrophages, microglial cells, astrocytes and oligodendrocytes, the ependymal epithelium, and the choroid plexus. Different strains of SRLV show varying neurotropism and varying neurovirulence, and the two attributes appear to be independent.
Lesions are not due to viral replication, but rather to the induction of an immune inflammatory response. Goats and sheep produce a strong humoral and cell-mediated immune response to SRLV infection. The host immune response likely plays a significant role in the development of lesions; experimentally, administration of immunosuppressive drugs almost abolished early lesions of visna in sheep. The host immune response is likely directed against virus-induced antigens rather than host antigens, and amplification of the immune response by infection results in a large influx of macrophages and lymphocytes and secretion of cytokines, which contribute to the damage of uninfected cells in the affected tissue, resulting in the lesions described below in the section on necropsy.
Infection with CAEV is lifelong. How and why the infection remains dormant for long and variable periods of time is not fully understood. For example, it is interesting that the neurologic form of CAE occurs more commonly in young animals, while visna in sheep is more common in adults. Strain differences and variations in neurotropism and neurovirulence as well as host factors may play a role.
Clinical Findings
In the neurologic form of CAE, the majority of affected animals are dairy breed kids between 1 and 6 months of age, though older goats also can be affected. The disease develops slowly over a period of weeks and is characterized by a progressive paresis and paralysis of the limbs, suggesting principally a spinal cord involvement. The hindlimbs are almost always affected, but tetraparesis is common and asymmetrical limb involvement may also occur. Kids may initially show some knuckling and inappropriate placement when standing and incoordination when walking, particularly in the hindlimbs (Figure 5.4a). Gradually, the animal may have increased difficulty rising (Figure 5.4b) and then be unable to rise at all.
Reflexes and muscle tone are likely to be increased, but are sometimes found to be decreased if the gray matter of the spinal cord is involved. When the hindlimbs are more affected than the forelimbs, which is common, the goat may pull itself around on the ground with the forelimbs. At least in the early stages of disease, many affected kids remain bright and alert and continue to eat and drink
Figure 5.4 Progressive paresis in a kid with the nervous form of caprine arthritis encephalitis virus infection. (a) Lack of proprioception manifested by cross - legged rear limb stance and knuckling of hind foot. (b) More advanced stage with inability to stand on hindlimbs. Source: Courtesy of Dr. Linda Collins Cork.
despite locomotor difficulties. Eventually, affected kids usually succumb to pneumonia, exposure, or other secondary disease problems if not euthanized first. Rarely, recoveries have been reported.
Clinical signs in the neurologic form of CAE are not always limited to locomotor deficits. In a retrospective of 30 cases, additional neurologic signs were observed in more than 50%. These signs included depression, blindness, abnormal pupillary response, nystagmus, opisthotonos, head tremor, head tilt, torticollis, circling, facial nerve deficits, and dysphagia. A variable increase in body temperature was also noted in the majority of cases (Norman and Smith 1983).
Clinical Pathology and Necropsy
Serology, using agar gel immunodiffusion (AGID) or ELISA tests, can be helpful in establishing the presence of retroviral infection. However, identifying antibody does not confirm that clinical disease is caused by retrovirus, because animals can be subclinically infected with CAEV yet have neurologic disease due to other causes. Conversely, serum antibody levels may fall below the sensitivity threshold of the AGID test, resulting in false-negative results. In addition, animals that were only recently infected may not yet have produced detectable antibodies. PCR assays on peripheral blood mononuclear cells from whole blood samples may be useful to identify the presence of viral antigen in suspect animals that are seronegative. Techniques for virus isolation from samples taken either from living animals (blood or milk leukocytes) or at necropsy (lung, choroid plexus, synovial membrane, or udder) are also available (OIE 2018c). Colorometric tests that can provide rapid results in the field are being developed using lateral flow technology (Tu et al. 2017) and loop-mediated isothermal amplification technology (Balbin et al. 2017).
The CSF may suggest a viral encephalomyelitis without indicating the definitive cause. Total protein content and white blood cell counts tend to be mildly to moderately increased. In most cases the increase in cellularity is caused by lymphocytes and monocytes, but increased neutrophils have also been observed. Median CSF total protein concentration in cases of CAE has been reported as 80 mg/dL, and median white blood cell counts as 26.5/mm3 (Smith 1982; Norman and Smith 1983). The hemogram is variable and of little diagnostic value.
At necropsy, gross lesions, when present, are limited to cloudiness of the meninges, focal brownish discoloration of the white matter in the brain and spinal cord and the ventricular surfaces, and/or swelling of the spinal cord. Microscopic lesions described in adult goats in Germany and Sweden initially presumed to have visna or visna-like disease (Stavrou et al. 1969; Dahme et al. 1973; Griem and Weinhold 1975; Sundquist et al. 1981) were similar to those described for the neurologic form of CAE (Cork et al. 1974; Norman and Smith, 1983). The lesions are similar to those originally described in sheep with visna (Sigurdsson et al. 1962).
Histologically, SRLV infections are characterized by a multifocal, mononuclear inflammatory leukoencephalo- myelitis accompanied by extensive demyelination. The inflammatory response is principally perivascular and is composed of aggregates of lymphocytes, macrophages, and plasma cells. In all cases, areas of inflammation are ringed by areas of increased astrocytes and microglia. Demyelination of nearby axons is a prominent finding. Axons are destroyed and there is general malacia in severely affected areas. In general, lesions are distributed periventricularly and periaqueductally and extend into the white matter along vessels. In severe cases, lesions may be found in adjacent gray matter. Lymphocytic infiltration of the meninges and the choroid plexus is a common finding.
Early reports of caprine visna and CAE suggested a possible difference in the two diseases based on location of lesions in the CNS, with CAE less likely to involve the brain and more likely to involve selected regions of the spinal cord (Cork et al. 1974). However, later studies of CAE demonstrated that brain lesions are common, occurring in as many as 60% of cases either with or without concurrent spinal cord lesions, and that spinal cord lesions are widely distributed and not often restricted to the cervical or lumbar regions (Norman and Smith 1983). Mononuclear inflammatory responses similar to those described in the CNS may also be found in the joints, lungs, and other tissues of goats with CAE.
Diagnosis
Currently the presumptive diagnosis of the neurologic form of CAE is based on a history of other forms of CAE in the herd, e.g., arthritis or mastitis, characteristic clinical signs, evidence of antibody in the affected goat, and typical neuropathology at the time of necropsy. Definitive diagnosis of any CAE infection depends on in vitro cultivation of virus from affected tissues.
The predominant sign in neurologic disease caused by CAEV is progressive paresis and paralysis. In young kids, the differential diagnosis must include enzootic ataxia due to copper deficiency, vertebral body or spinal cord abscesses, congenital abnormalities of the spinal cord or vertebral column, and cerebrospinal nematodiasis. If the onset is reported to be acute because early signs of paresis were overlooked, then enzootic muscular dystrophy, spinal trauma, tick paralysis, and polyradiculoneuritis also must be ruled out. Multiple conditions may be concurrent, as in the reported cases of CAE and enzootic ataxia occurring in goat kids in the northeastern United States (Norman and Smith 1983; Lofstedt et al. 1988).
In adult goats, cerebrospinal nematodiasis, abscesses, possible delayed organophosphate neurotoxicosis, and scrapie must be considered in the differential diagnosis. Unlike in sheep, intense pruritus is less commonly reported in caprine scrapie and therefore the condition may look more like an SRLV infection than it would in sheep.
When lesions suggestive of brain involvement are observed, PEM and listeriosis should then also be considered in the differential diagnosis. In all cases rabies should be considered, because it can also present as an ascending paralysis.
Treatment and Control
There are no known treatments for retroviral infections of small ruminants in veterinary medicine. In the case of
CAE, the control methods described in Chapter 4 for the arthritic form of the disease also apply to the neurologic form.
Border Disease
Border disease, also known as “hairy shaker” disease in lambs, is an infectious, contagious viral disease of sheep and goats. It causes abortions, infertility, stillbirths, weak lambs and kids, and a characteristic pattern of tremors, or shaking, in newborn lambs and kids. In lambs, it also produces a characteristic hairy appearance to the fleece and histologic changes in the skin. These skin changes are not seen in affected kids. More details about border disease in sheep are available elsewhere (Constable et al. 2017).
Etiology
Border disease is caused by a pestivirus. The pestiviruses are enveloped, single-stranded, positive-sense RNA viruses in the family Togaviridae. The pestivirus genus historically has included four related species responsible for livestock diseases: border disease virus (BDV), bovine virus diarrhea virus-1 (BVDV-1), bovine virus diarrhea virus-2 (BVDV-2), and classical swine fever (hog cholera) virus (CSFV). Tlie viruses share physical, chemical, and biologic properties, including a shared common soluble antigen. Cross-species infections are known to occur, as antibodies against pestivi- rus have been identified in more than 40 ruminant species (Hamblin and Hedger 1979). BDV can infect cattle and swine in addition to sheep and goats, while BVDV-1 and BVDV-2 can infect goats, sheep, and swine in addition to cattle.
Definitive classification of the pestiviruses is currently a work in progress. Recent phylogenetic analysis of the pes- tiviruses based on the Npro gene encoding for the non- structural protein Npro indicates that these viruses do not indeed segregate according to their traditional classifications based on host species origin (e.g., bovine versus porcine versus ovine; Becher et al. 1997). Based on this analysis, three pestivirus isolates derived from goats were classified in the BVDV-1 genotype rather than the BDV genotype traditionally associated with border disease in sheep and goats. BVDV-1 was subsequently isolated from goat kids from mixed sheep and goat flocks in Italy with signs consistent with border disease, while BVDV-2 was identified in sheep from those flocks (Pratelli et al. 2001). An isolate from goats with border disease-type signs in the Republic of Korea was classified in the BVDV-2 genotype, again on the basis of phylogenetic analysis of the Npro gene (Kim et al. 2006). It is now clear that BDV can infect cattle and swine in addition to sheep and goats, while BVDV-1 and BVDV-2 can infect goats, sheep, other wild ruminants, and swine in addition to cattle.
Additional phylogenetic analysis of pestivirus isolates indicated that the BDV genotype could be further divided into four subgroups: BDV-1, BDV-2, BDV-3, and BDV-4 (Becher et al. 2003; Arnal et al. 2004). Subsequent to that, a pestivirus isolated from caprine fetuses in a mixed sheep and goat herd in central Italy experiencing abortions was characterized phylogenetically as a BDV, but, based on its Npro sequence, could not be included in one of the existing subgroups. Therefore, an additional new BDV subgroup was proposed (De Mia et al. 2005). Since then, additional genomic studies have proposed as many as 9 (Giangaspero 2011) to 11 or more species (Smith et al. 2017) of pestivirus, with at least 4 or possibly 5 of them that infect goats and sheep and can produce clinical disease consistent with a diagnosis of border disease.
Epidemiology
The incidence of border disease in goats and its impact on the goat industry are unclear. The disease in sheep occurs in North America, Europe, China, Australia, and New Zealand. In contrast, naturally occurring, clinical border disease in goat kids has been reported only sporadically. While the disease was first described in sheep in 1959, in the border region between England and Wales, the first naturally occurring case of border disease in a goat herd was not reported until 1982, from Norway (Loken et al. 1982b). However, there was a report of a pestivirus isolated from the lungs of a 4-week-old kid with pneumonia in Australia the previous year (Fraser et al. 1981). Since then there has been little additional documentation of clinical border disease in goat flocks, save for some reports of goat cases of abortion and stillbirths along with sheep cases in mixed flocks in Italy (Pratelli et al. 1999; De Mia et al. 2005). These goat-related cases may have involved BVDV virus as well as BDV virus. BDV has been reported as a cause of diarrhea in several goat herds in two provinces of eastern China, with morbidity up to 37% and mortality up to 15% in some affected herds (Li et al. 2013).
Despite the relative lack of clinical reports, serologic surveys performed on goats in Canada (Elazhary et al. 1984; Lamontagne and Roy 1984), the United States (Fulton et al. 1982), Austria (Krametter-Frotscher et al. 2007), Norway (Loken 1989), Nigeria (Taylor et al. 1977), Chile (Celedon et al. 2001), Brazil (Flores et al. 2005), and Turkey (Alpay et al. 2018) indicate the presence of antibodies to BVDV or BDV in the range of 3-16% of goats tested. The significance of antibody responses in goats is not clear. They may merely represent an exposure to sheep with BDV, cattle with BVDV, or actual caprine infection. It is clear at least that the exposure of goats to pestivirus has a wide geographic distribution.
It is possible that clinical border disease in goats is frequently unrecognized because of a different disease pattern in goats than in sheep. In experimental caprine infection, early abortion, fetal resorption, and mummification occur more commonly than in sheep, while the delivery of live, weak term kids, or kids with tremors, occurs less commonly. In a New Zealand study, border disease was implicated or suspected in 23% of caprine abortions (Orr et al. 1987; Orr 1988). In an episode of naturally occurring caprine border disease in Norway, only one shaker kid was identified, but 43% of does in the herd were seropositive, a considerably higher percentage than observed in general field surveys (Loken et al. 1982b). An outbreak of border disease affecting five goat herds was reported from Norway in which the source of infection was an orf vaccine contaminated with pestivirus (Loken et al. 1991). There was notable reproductive failure, with barrenness, abortion, and birth of weak or dead kids, but no live offspring showed characteristic signs of border disease, though lambs present in one flock did show neurologic signs.
Pathogenesis
Horizontal and vertical transmission of the virus occur. Horizontal transmission is believed to be by ingestion or aerosol inhalation of virus. BDV has been isolated from sheep keds (Melophagus ovinus) in China, but their significance in disease transmission is unclear (Liu et al. 2019).
The fetus is infected transplacentally in vertical trans - mission. The outcome of intrauterine and fetal infection depends to a great extent on the time of infection, although virus strain variation and breed factors are believed also to play roles. In an experimental challenge of goats, infection at 40 days of gestation resulted in no live kids born, infection at 60 days resulted in 11% live kids born, and infection at 100 days in 73% live kids born (Loken and Bjerkas 1991). When fetuses survive an infection that occurs before the onset of immune competence, they are born with a persistent infection that is probably lifelong. If they are bred, their own offspring will be persistently infected as well.
Intrauterine infection produces a more severe placentitis in goats than in sheep, and this may account for more frequent fetal death and less frequent delivery of full-term shaker kids when compared with sheep (Barlow et al. 1975; Huck 1973). Infection of fetuses in utero can result in the birth of immunotolerant kids if infection occurs as late as 100 days of gestation. Healthy-appearing kids as well as shaker kids born to infected does can carry and shed the virus, serving as a source of new infections (Loken and Bjerkas 1991).
Goat fetuses and kids from experimentally infected does all show some characteristic CNS lesions, regardless of whether they show clinical signs of tremors after birth. Goat kids, however, do not have the characteristic dermal follicular changes associated with the hairy fleece in affected lambs (Orr and Barlow 1978).
Clinical Findings
Shaker kids and lambs show muscle tremors right from birth. These rhythmic tremors are most prominent in the hindquarters, but may extend up the trunk and neck. The general impression is one of jerkiness. Affected young have difficulty rising and when they do they have an awkward gait. Nursing is difficult and affected young may become quickly hypothermic and hypoglycemic. They appear weak, listless, and depressed if not attended to. The long bones of the limbs may feel finer than usual, and the head may appear narrowed and exaggeratedly convex in the frontal area. No abnormalities of the haircoat or skin occur in affected goats (Orr and Barlow 1978). Does show no clinical signs other than a history of barrenness, abortion, stillbirth, or fetal mummification.
It is possible that clinical manifestations may vary depending on the specific pestivirus species or genotype infecting goats. For example, a border disease-like syndrome occurring in the Republic of Korea since 1998 included diarrhea and a high mortality rate, in addition to the expected signs of abortions, stillbirths, and kids with neurologic signs. Laboratory investigations revealed that the causative agent was BVDV-2 (Kim et al. 2006).
Clinical Pathology and Necropsy
Seroconversion against pestivirus antigens in association with abortion in a herd is suggestive of border disease. ELISA and VN tests are available for quantifying antibody responses. In the one reported field outbreak in Norway, 90% of does with serum-neutralizing antibodies to the BVD virus had titers of 1 : 500 or more (Loken et al. 1982b). In affected kids, absence of antibody does not rule out border disease, due to the possible development of in utero immunotolerance. In such kids, infection can be confirmed by testing for the presence of viral antigen or nucleic acid in blood samples (OIE 2018a).
For definitive diagnosis of border disease in abortion cases, it is recommended that formalin-fixed fetal brain, fresh fetal kidney and liver, and maternal serum be submitted for histology, virus isolation, and serology (Orr 1988). In addition to virus isolation, ELISA techniques are available for the detection of BDV antigen. RT-PCR and in situ hybridization are available to detect nucleic acid of BDV (OIE 2018a). As goats and sheep may be infected with BVDV as well as BDV, it is preferable to use diagnostic assays that are broadly reactive and will readily detect all strains of BDV and BVDV.
No gross lesions are observed at necropsy of affected kids. Histologic lesions in the CNS are characterized by pronounced hypomyelinogenesis, hypergliosis, and vasculitis, particularly in the white matter of the cerebrum and cerebellum. There is perivascular gliosis and infiltration of vessel walls with lymphocytes and histiocytes. Corpora amylacea are also noted. In aborting does, there is a marked placentitis with necrotizing carunculitis. Virus may be isolated from uterine contents or vaginal discharges. Fetuses may be autolyzed, mummified, or grossly normal, but may show reduced numbers of myelinated fibers in the ventral spinal tracts histologically.
Diagnosis
Diagnosis is based on a history of reproductive failure, occurrence of newborn shaker kids, and laboratory confirmation of infection. The differential diagnoses for reproductive failure and abortion are discussed in Chapter 13. Hypoglycemia, septicemia, or meningitis may produce convulsions in newborn kids that could be mistaken for border disease shakers. When kids have difficulty rising, the following should be considered: nutritional muscular dystrophy, congenital vertebral malformations, spinal trauma, and swayback.
Treatment
There is no specific treatment. Supportive care to ensure regular feeding promotes kid survival, but the overall herd status must be considered with regard to the advisability of keeping potential carrier animals in the herd.
Control
All does that abort, regardless of cause, should be isolated from the herd. The BDV is present in vaginal discharges after abortion or kidding. In general, goats should be maintained separately from cattle and sheep. Does producing shaker kids should be culled, on the presumption that they themselves are infected. Elimination of infection from the herd by identification and culling of persistently infected individuals before they reach breeding age is advised. However, identifying all persistently infected animals by serologic testing can be problematic, because some nonreactors may in fact be persistently infected, immunotoler- ant shedders of the virus, and tests to detect viral antigen or nucleic acid may be required. Herds and flocks free of border disease should be maintained as closed herds and flocks without the purchase of animals from outside sources.
While the use of experimental inactivated BDV vaccines has been reported (Brun et al. 1993), there are no such vaccines commercially available. The use of BVDV vaccine has been suggested in sheep flocks suffering annual outbreaks of border disease, but this pattern of disease has not been reported in goat herds. If vaccines are used, vaccine failures may occur, because different strains of BVDV and BDV virus may not be cross-protective. However, a field study in France indicated that vaccination of sheep with a half dose of commercial inactivated BVD-1 vaccine marketed for cattle produced neutralizing antibodies in sheep against BVD-1 as well as against the two different BDV genotypes that were circulating in sheep flocks in the region (Anne 2012).
Louping-Ill and Related Infections in Small Ruminants
Louping-ill, also known as ovine encephalomyelitis, is an acute, non-contagious encephalomyelitis caused by the louping-ill virus (LIV), an enveloped, positive-sense, single-stranded RNA virus in the genus Flavivirus. LIV is included in the tick-borne encephalitis virus complex (TBEV) that also includes Kyasanur Forest disease virus and Alkhurma virus. Phylogenetic studies have been conducted on LIV (Gao et al. 1997; McGuire et al. 1998). The disease is documented to occur only in Scotland, England, Wales, Ireland, and Norway. There is also evidence of infection and illness of sheep and goats, with several additional flaviviruses related to but genomically distinct from LIV that have been recognized in Europe: the Greek goat encephalomyelitis virus (Papa et al. 2008), the Spanish goat encephalomyelitis virus (SGEV) (Mansfield et al. 2015), the Turkish sheep encephalitis virus (TSEV) (Gao et al. 1993), and the Spanish sheep encephalomyelitis virus (SSEV) (Gould et al. 2003).
The LIV is maintained primarily in populations of sheep, red grouse, and mountain hares on tick-infested moors or pastures. It is transmitted by the three-host tick vector Ixodes ricinus, which can transfer the virus transstadially but not transovarially. The virus may also be transferred by blood-contaminated needles, instruments, and contact with infected tissues.
Clinical disease, manifested mainly as an encephalomyelitis, occurs most commonly in sheep, but other animals are sporadically reported to be affected, including goats, dogs, horses, cattle, pigs, farmed red deer, and llamas (Macaldowie et al. 2005). Louping-ill has serious zoonotic potential: the virus can produce fatal meningoencephalitis in humans. Because of the zoonotic risk, veterinarians and producers in endemic areas should observe proper safety precautions when handing sick animals or carcasses with signs suggestive of louping-ill.
In sheep, the clinical signs include fever during the initial viremic stage, followed by neurologic signs once the virus enters the CNS. These include fine muscle tremors, ataxia, nibbling motions, weakness, and collapse, with death occurring one to three days following onset of signs. During the course of the disease, the sheep may walk with jerky, stiff, almost bouncy movements, which account for the name louping-ill. There is one report of suspected louping-ill occurring in a goat recently arrived on the Scottish island of Islay (Gray et al. 1988). The suspect goat showed fever, trembling, forelimb weakness, and retching, became recumbent, and died. It had a high reciprocal hemagglutination inhibition antibody titer to the LIV and had characteristic histologic lesions in the brain.
A provisional diagnosis of louping-ill was made in an outbreak of neurologic disease in a herd of 70 goats in northern Spain in 2011, which had recently been introduced into the area from southern Spain (Balseiro et al. 2012). The first goat to become ill initially showed hindleg lameness, rapidly followed by incoordination, fever, tremors, bulging eyes, and death two days later. Over the following four months an additional 17 goats showed the same signs and died. Heavy tick infestations were noted on affected animals. Subsequent phylogenetic analysis of the virus involved indicated that the causative agent was in fact SGEV and not LIV (Mansfield et al. 2015). There were no gross lesions in necropsied goats and histologic lesions were limited to the CNS. In the cerebellum there was necrosis of Purkinje cells and neurons. Other lesions were concentrated in the hypothalamus and midbrain as well as the medulla oblongata and spinal cord, and consisted of extensive lymphocytic perivascular cuffing as well as neu- rophagia and gliosis, characterized by degenerating neurons surrounded by glia cells. The lesions were indistinguishable from those observed in louping-ill. Goats can be experimentally infected with LIV, but appear to be more resistant than sheep (Reid et al. 1984). All challenged does became viremic and shed the virus in their milk for up to nine days after challenge. This resulted in infection of suckling kids with more severe clinical disease. These findings suggest that subclinically infected dairy goats could potentially transmit louping-ill to people consuming unpasteurized milk.
Confirmation of the disease requires histologic examination of the brain, virus isolation from CNS tissue, and serology using serum neutralization or hemagglutination inhibition assays. Numerous molecular techniques are now available for identification of the various flaviviruses affecting small ruminants, notably RT-PCR for detection of LIV (Marriott et al. 2006) and for SGEV (del Cerro et al. 2018).
There are no specific treatments, but some affected animals with LIV may recover with nursing and supportive care. An inactivated tissue culture vaccine, which can be given to sheep, goats, and cattle, is in use in the United Kingdom. A single dose should be administered four weeks before turnout to tick-infested pastures. Immunity is considered to last for at least two years. The commercial louping-ill vaccine available in the United Kingdom has been shown also to protect goats against experimental challenge with the Spanish goat encephalitis virus (Salinas 2017).
Borna Disease
Borna disease is a non-suppurative viral polioencephalomyelitis that historically has affected horses and sheep in central Europe. Recent studies suggest that the virus is more widely distributed than previously thought, that species other than horses and sheep may be infected, and that the disease also may be an emerging zoonosis, though the latter point remains controversial. Goats can be infected experimentally (Ihlenburg 1962) and there is evidence that naturally occurring caprine cases have occurred, though rarely. Several reviews of progress in the understanding of Borna disease and the controversies surrounding it have been published (Ludwig and Bode 2000; Staeheli et al. 2000; Dauphin et al. 2002; Durrwald et al. 2006).
Etiology
The causative virus is now identified as Borna disease virus (BoDV) in the newly created family Bornaviridae in the order Mononegavirales. It is an enveloped, single-stranded, negative-sense RNA virus measuring 100-130 nm in diameter. Several unique aspects of the virus led to the creation of the new Bornaviridae family, as summarized by Dauphin et al. (2002). BoDV is the only negative, non-segmented, single-stranded RNA animal virus with a nuclear site of replication and transcription. Its genome compaction is overcome by the overlapping of ORF and transcription units and by posttranscriptional RNA splicing. The virus is characterized by strict neurotropism. It is non-cytolytic and has a low rate of replication and persistence in the CNS. Unlike the majority of RNA viruses, the BoDV genome sequence is extremely stable over time.
Epidemiology
The disease has been known for almost two centuries in horses in southwestern Germany and the name is associated with the town of Borna in Saxony, where the disease later emerged and peaked in the 1890s. The disease is also recognized to affect sheep. Reports of the disease in those two species remained localized to Germany for most of the history of the disease, until it spread through the Rhine valley to Switzerland, Austria, and Liechtenstein starting in the 1970s. In this enzootic region of central Europe, horses and sheep remain the principal species affected, but there have also been confirmations in small numbers of cattle, donkeys, dogs, deer, rabbits, South American camelids, and goats. The goat cases were recorded in Switzerland, two from the same premises in 1987 and a single case in 1995 (Caplazi et al. 1999). The diagnoses were retrospective by IHC. No clinical descriptions were provided.
In the 1960s, there were reports of a neurologic condition in the Middle East. It was first described as near east equine encephalitis, but then later proposed as Borna disease based on clinical and histopathologic similarities, including the presence of Joest-Degen bodies in the brains of affected horses, a characteristic of Borna disease (Daubney 1967). Two cases of putative Borna disease in goats in Lebanon were included in those reports (Daubney 1967). It now appears unlikely that these were truly cases of Borna disease (Rott et al. 2004).
Contemporary understanding of Borna disease suggests that the cases described were caused by a virus other than BoDV, because BoDV does not produce a cytopathic effect in cell culture. Moreover, the lesion profile described is not consistent with the histopathology of classical Borna disease, and the inclusion bodies initially identified as Joest- Degen bodies may in fact have been multiple nucleoli, which occur quite often in large neurons (Prof. Dr. Felix Ehrensperger, Department of Veterinary Pathology, University of Zurich, Zurich, Switzerland, personal communication, 2007).
BoDV has been reported to be responsible for disease occurrences in ostriches in Israel (Malkinson et al. 1993), cats in Sweden (Lundgren and Ludwig 1993), and horses in Japan (Taniyama et al. 2001) and Iran (Bahmani et al. 1996). The virus appears to have a much more cosmopolitan distribution that goes beyond the enzootic region of central Europe. In 2006, subclinical infections were reported in Chongqing goats in China based on detection of the BoDV p24 gene in brain tissue and peripheral blood mononuclear cells (Zhao et al. 2006).
Borna disease has also been considered as a possible cause of neurologic and psychiatric disease in humans in various countries including the United States, Japan, and Germany (Rott et al. 1991). The role of BoDV as a zoonotic agent has been controversial and the evidence for and against has been reviewed (Chalmers et al. 2005; Durrwald et al. 2007). However, eight cases of human encephalitis occurring in Germany between 1999 and 2019 have now been definitively confirmed as being due to BoDV infection based on detection of BoDV RNA and BoDV-reactive antibodies as well as isolation of the virus from one of the affected patients, thus confirming that the disease is zoonotic (Niller et al. 2020).
In the enzootic region of central Europe, Borna disease historically has a seasonal occurrence, with a peak in the spring. This has led to the presumption that an agent reservoir exists, most likely in wildlife populations, though considerable efforts have been undertaken to identify such reservoirs, without definitive success. The bicolored, white-toothed shrew (Crocidura leucodon) is considered to be the agent reservoir (Hilbe et al. 2006).
Knowledge of the transmission of Borna virus is incomplete. The virus is shed in nasal, salivary, and conjunctival secretions, and inhalation and ingestion are considered to be important routes of transmission. Viral RNA and proteins are also found in peripheral blood mononuclear cells, so blood-borne infection may be possible. However, some efforts to demonstrate horizontal transmission from horse to horse or sheep to sheep have been unsuccessful (Staeheli et al. 2000). Infected animals develop an antibody response, but it is not protective. The role of carrier animals in transmission remains unclear.
Clinical Findings
Clinical findings in goats are not well documented. In other species the clinical presentation reflects neurologic disease, but specific signs may vary between affected animals and include behavioral changes, paresthesia, motor dysfunction, incoordination, paralysis, and death. In sheep, disturbances of sensory function are more pro - nounced than signs of motor dysfunction, which are more predominant in horses. However, head pressing, staggering, and ataxia are reported in sheep as the clinical course progresses over a period of 4-10 days (Ludwig and Bode 2000).
In the putative cases of caprine Borna disease reported from Lebanon, the clinical findings included fever, anxiety, bleating, salivation, champing of the jaws, partial facial paralysis, fine tremors, head pressing, circling, paraplegia, convulsions, and death (Daubney 1967).
Clinical Pathology and Necropsy
Serologic tests can be applied to blood or CSF. Western blot, ELISA, and immunofluorescence assay (IFA) have been applied, with the latter considered to be the most sensitive. Antibody levels may be low in acute clinical cases of Borna disease, so serologic tests must have a high sensitivity. Antibodies may be undetectable in subacute and chronic cases (Dauphin et al. 2002). For these reasons, efforts to identify virus or viral antigen should also be pursued.
Virus isolation in cell culture can be attempted from the brain tissues of affected animals, but the number of infectious particles may be low, leading to false-negative results. IHC may be more rewarding. RT-PCR is now being used to detect virus in blood and brain tissue.
Gross lesions are non-specific and may include mild leptomeningitis, congestion of cerebral vessels, and hemorrhage between the cerebrum and cerebellum. Histologic changes are consistent with viral encephalitis and are characterized by perivascular and parenchymal infiltration of lymphocytes in affected portions of the brain. Characteristic inclusion bodies, known as Joest-Degen bodies, may be present in the nuclei of infected neurons, but their presence is variable.
Diagnosis
A presumptive diagnosis may be made on the basis of signs of encephalomyelitis in goats in countries or regions where Borna disease is enzootic, with the realization that the disease is quite rare in goats and that there are more common causes of caprine encephalomyelitis. Definitive diagnosis is by laboratory confirmation. Other possible causes of acute encephalitic signs in goats include rabies, pseudorabies, PEM, bacterial meningoencephalitis, and possibly CAE.
Treatment and Control
There is no treatment for Borna disease and the prognosis for recovery is guarded. In species other than goats, affected animals sometimes recover spontaneously but then relapse. The disease does not occur commonly in goats and specific control measures directed toward goats are not formulated. Given the zoonotic potential of Borna disease and other conditions such as rabies that may appear like Borna disease, proper precautions should be taken when handling clinical cases or during necropsy.
West Nile Encephalomyelitis
West Nile encephalomyelitis is caused by West Nile virus (WNV), a mosquito-borne flavivirus. WNV can infect numerous species of mammals, birds, and even some reptiles, but causes clinical, neurologic disease, mainly in horses and humans. To date there are no confirmed cases of the disease in goats from anywhere in the world, though antibodies to WNV are readily detectable in goats in those regions where WNV is present and circulating through mosquito populations. This indicates that goats can be infected with WNV following the bite of mosquitoes and that they mount an immune response, but their likelihood of developing clinical disease is very low. Therefore, a diagnosis of West Nile encephalomyelitis should not be made in a goat with neurologic disease based solely on the presence of antibody to WNV in a single serum sample. A fourfold rise in titer between acute and convalescent serum samples taken two weeks apart and initiated at the onset of disease would be far more convincing, but other causes of neurologic disease in goats should nevertheless be ruled out.
Etiology and Epidemiology
WNV is a single-stranded RNA virus in the genus Flavivirus and is a member of the Japanese encephalitis virus sero- complex. Members of this group of flaviviruses have a close antigenic relationship, which accounts for serologic crossreactions that occur in laboratory diagnosis. These viruses include WNV, Japanese encephalitis, St. Louis encephalitis, Murray Valley encephalitis, and Kunjin virus (Petersen and Marfin 2002).
WNV is an arbovirus and is maintained in a primary enzootic cycle involving Culex spp. mosquitoes and wild birds of many species. Migratory birds may carry the infection to new locations and infection may become established in those locations if there are suitable mosquito hosts present. In temperate regions, mosquitoes hatch in the late spring and feed on birds through the spring and summer, so that the virus population in birds and mosquitoes is much amplified by late summer/early autumn. At that time, certain populations of mosquitoes that serve as “bridge vectors” also feed on humans and horses and other mammals, as well as on birds, creating a secondary cycle that can lead to clinical disease in humans and horses and possibly other animals. Humans, horses, and other mammals, however, do not produce sufficient viremia in response to WNV infection to pass the virus back to mosquitoes, and are therefore dead-end hosts. The primary bird/mosquito cycle is required to maintain the WNV reservoir.
The name of the virus derives from the fact that it was first isolated from a woman with fever in the West Nile district of Uganda in 1937. The first documented disease epidemic in humans associated with WNV occurred in Israel in 1951 (Bernkopf et al. 1953). The virus became recognized or established in many countries of Africa, the Middle East, western Asia, and southern Europe in subsequent years, affecting horses as well as humans. In 1999 the disease was introduced to North America, with the first cases recognized in birds in New York City. By 2005, the virus was present in Culex spp. mosquitoes and/or wild bird populations in all 48 of the continental US states and most states had reported human and/or equine cases. The disease is now widespread in Mexico and Canada as well. It is not clear how the disease was introduced into North America.
Among the domestic animals, horses and other equids are the most likely to exhibit clinical disease following infection, and among domestic poultry, geese and very young chickens are most likely to become ill. Farmed alligators have been definitively diagnosed with West Nile encephalitis in Georgia and Florida (Miller et al. 2003; Jacobson et al. 2005). Clinical illness in ruminants is rare and experimental infection of ruminants is not consistently achieved (McLean et al. 2002). There have been no reported cases in goats, even from countries such as Israel, which has a reasonably large goat population and where the virus has been endemic for more than 50 years (Dr. Shmuel Zamir, Chief Sheep and Goat Health Officer, Veterinary Services and Animal Health, Ministry of Agriculture and Rural Development, Government of Israel, personal communication, 2007). A case of West Nile encephalitis in a 6-month-old sheep from Nebraska was confirmed by RT-PCR assay in 2002 (Callan and Van Metre 2004) and a second 2-year-old sheep from Missouri was confirmed with West Nile encephalitis by similar methods the same year (Tyler et al. 2003). In camelids, 8 of 17 alpacas demonstrating neurologic signs were confirmed as having West Nile encephalitis by PCR in Colorado in 2003 (Callan and Van Metre 2004).
While there are no reported clinical cases in goats, there are numerous reports of goats with circulating antibody to WNV. Countries where serologic surveys have revealed WNV antibody-positive goats include Romania with 3.2% of goats sampled testing positive (Topciu et al. 1971), Greece with 8.7% (Koptopoulos and Papadopoulos 1980), Pakistan with less than 5% (Go 1990), India with 6.75% (Mall et al. 1995), Nigeria with 18% (Olaleye et al. 1990), and Turkey with 2.85% (Albayrak and Ozan 2013). Antibody to WNV has also been identified in goats in the United States (Dr. Richard Bowen, Professor, Department of Biomedical Sciences, Animal Reproduction and Biotechnology Laboratory, Colorado State University, Fort Collins, CO, personal communication, 2007). This data underscores the fact that goats are regularly exposed to WNV in enzootic countries. The lack of clinical case reports suggests either that they are resistant to infection or that the low-level viremia associated with ruminant species is unlikely to result in clinical manifestation of disease. There are concerns, however, that more virulent strains of WNV could emerge and the disease patterns in mammalian species could change in the future as a result (McLean et al. 2002).
Clinical Findings
Because there are no reported confirmed cases in goats, no reliable clinical findings can be reported for this species. In humans, the most common presenting signs are fever, weakness, nausea, vomiting, and changes in mental status or confusion (Petersen and Marfin 2002). In horses, a retrospective study of 569 cases indicated the following signs present, in decreasing order of frequency: incoordination, muscle tremors, twitching of the face or muzzle, weakness or paralysis of limbs, caudal paresis, recumbency or difficulty rising, lip droop, teeth grinding, fever, circling, and blindness. Of the 569 horses, 345 (61%) recovered, 126 (22%) died, and 98 (17%) had an unknown outcome (Schuler et al. 2004).
In the two reported cases in sheep, both animals presented in recumbency of five to seven days' duration. One sheep showed muscle fasciculation as well as hyperesthesia, with brief muscle spasms and extensor rigidity when touched. It also exhibited tonic clonic convulsions of short duration on presentation. These convulsions became more frequent and severe and the animal was euthanized (Tyler et al. 2003). The second sheep was mentally depressed, unable to stand, and exhibited spontaneous nystagmus as well as muscle fasciculation. It too was euthanized (Callan and Van Metre 2004).
Clinical Pathology and Necropsy
There are no reports of laboratory or necropsy findings in goats with WNV encephalitis. In other species, CSF analysis may reveal an elevated protein and increased cellularity comprising lymphocytes and large mononuclear cells.
At necropsy, in other species gross lesions in the CNS are limited to focal hemorrhage in the brain or spinal cord on cut section. Histologically, there is mild to moderate, multifocal, lymphocytic polioencephalomyelitis with perivascular cuffing, microgliosis, and some neuronal degeneration in severely affected areas (Cantile et al. 2001).
Diagnosis
In horses, the most commonly affected domestic mammal, diagnosis of WNV is currently based on observation of compatible clinical signs such as ataxia, paresis, paralysis, hyperesthesia, muscle fasciculation, seizures, or fever, and on one or more of the following: virus isolation or RT-PCR detection of WNV from tissue, blood, or CSF; a fourfold increase in plaque reduction neutralization test (PRNT) antibody titers between paired serum samples taken two weeks apart (in temporal association with clinical signs of disease); detection of immunoglobulin (Ig)M antibody to WNV by IgM-capture ELISA; or a neutralizing titer of more than 1 : 10 by PRNT in a single serum sample (Kleiboeker et al. 2004).
As the intensity of viremia and the strength of the antibody response may differ in goats relative to horses, it is not clear that these same diagnostic criteria would apply to goats. The most reliable criterion for goats would likely be documentation of a fourfold rise in titer between two serum samples, one taken at the start of clinical disease and the other 14 days later.
A necropsy should be performed with histopathologic examination of the CNS in goats that die with neurologic signs, as much to rule out other important diseases as to rule in WNV infection. The differential diagnosis should at least include rabies, pseudorabies, and listeriosis in any goat; pregnancy toxemia in pregnant females; and CAE in young goats. Depending on geographic location, other potential causes of encephalomyelitis, e.g., louping-ill, border disease, or Borna disease, should be considered. If other conditions are ruled out and WNV is still under consideration, then IHC and/or PCR techniques on brain and spinal cord should be conducted to identify WNV antigen.
Treatment and Prevention
There is no specific treatment for WNV encephalitis. Affected animals may recover, so efforts to provide supportive and nursing care are justified to ensure proper nutrition and hydration and to prevent injuries associated with incoordination or prolonged recumbency. Efforts to control mosquitoes around goats and goat barns could be considered. Commercial WNV vaccines are available for use in horses, but they are not approved for use in goats or other ruminants. Given the absence of reports of confirmed clinical disease in goats, there is no apparent economic justification for the use of WNV vaccines in goats.