Taxonomic Developments in Sporothrix
Sporothrix has great phenotypic plasticity. Phenotypic variations include changes in colony morphology, growth on different culture media, thermal tolerance, virulence in a murine model, susceptibility to antifungal drugs, protein secretion, genome architecture, and melanin synthesis (Marimon et al.
2008b; Fernandes et al. 2009a; Arrillaga-Moncrieff et al. 2009; Sasaki et al. 2014; Almeida-Paes et al. 2015; Della Terra et al. 2017). Recent phylogenetic studies have shown that S. schenckii is actually a complex of distinct species, with more than one taxon capable of causing disease in humans and animals (Zhang et al. 2015).The first molecular evidence that the classical agent S. schenckii is represented by two or more species came from the studies of de Beer et al. (2003) through sequencing and phylogenetic analysis of the ITS (internal transcribed spacer) region located in the ribosomal operon (rDNA). Strains of patients from the Americas and South Africa were classified in group I. Interestingly, the strains isolated directly from environmental sources such as soil and decaying plant material were classified in group II, with very few representatives of clinical origin. Thus, the phylogenetic splitting observed was consistent with the niche of these groups, separating clinical from environmental isolates (Fig. 10.2). In fact, Sporothrix samples from soil and decaying organic matter tend to be less pathogenic to murine models than clinical isolates (Howard and Orr 1963; Arrillaga-Moncrieff et al. 2009; Rodrigues et al. 2016a).
A few years later, Marimon et al. (2006) proposed three sibling species of medical interest within S. schenckii: Sporothrix brasiliensis, S. mexicana, and S. globosa. Marimon et al. (2007) also studied growth at different temperatures (30, 35, and 37 °C), assimilation of sugars (sucrose, raffinose, and ribitol), morphology of sessile conidia, and partial sequence of the calmodulin gene.
The analysis based on the calmodulin gene grouped the Sporothrix species in different clades: clade I comprised only Brazilian isolates (later named S. brasiliensis); clade II comprised most of the isolates from the USA and isolates from South America (later named S. schenckii s. str.); clade III included isolates from China, India, Italy, Japan, Spain, and the USA (later named S. globosa); and clade IV consisted of environmental isolates from Mexico (later named S. mexicana).Isolated in 1956, S. schenckii var. luriei was described by Ajello and Kaplan (1969) from a human in South Africa. This isolate exhibited morphological characteristics different from other S. schenckii isolates, including the presence of large, round cells with thick walls and typically with one septum, and the production of sclerotia in vitro when grown on oatmeal agar. Phylogenetic data demonstrated a clear separation between S. schenckii var. luriei (clade VI) and the other species in the pathogenic clade. Moreover, morphological and physiological characteristics
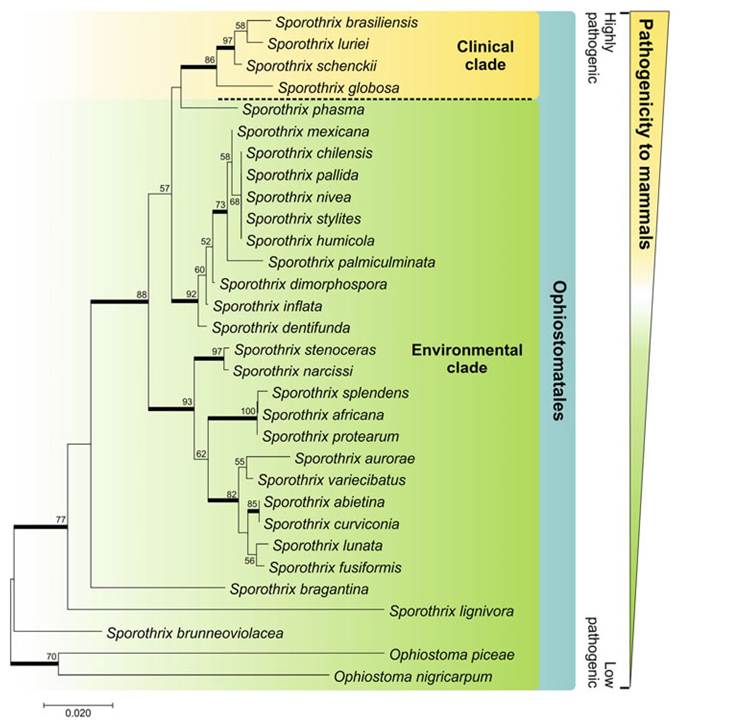
Fig. 10.2 Phylogenetic relations between members of clinical and environmental relevance in Sporothrix, based on sequences from the ribosomal operon rDNA (IT1 + 5.8 s + ITS2). Analysis method: neighbor joining (model: T93 + G). The numbers next to the branches refer to the percentages of resampling (1000 bootstraps)
showed differences between the species, including the absence of pigmented sessile conidia, presence of long conidia, good growth at 35-37 °C, and the inability to assimilate sucrose and raffinose; consequently, S. schenckii var. lurid was named as S. luriei (Marimon et al. 2008a).
Today, the “clinical clade” of Sporothrix is composed of S. brasiliensis (clade I), S. schenckii s. str. (clades IIa and IIb), S. globosa (clade III), and S. luriei (clade VI) (Zhou et al. 2014). The strains from human and animal sources are located in the clades S.
brasiliensis, S. schenckii s. str., S. globosa, and S. luriei. Located at a relatively large genetic distance, the environmental species of Sporothrix are flanked by environmental species of Ophiostoma (Fig. 10.2).The Sporothrix of clinical interest, often infecting vertebrate hosts, form a monophyletic group (Marimon et al. 2007; Roets et al. 2008); however, other genetically distant species within the Ophiostomatales can also be accidental agents of human and animal sporotrichosis (Fig. 10.2). This is the case of S. pallida, S. mexicana, S. chilensis, S. stenoceras, and Ophiostoma piceae (Bommer et al. 2009; Morrison et al. 2013; Rodrigues et al. 2013a; Scheufen et al. 2015).
The dichotomous key in Fig. 10.3 was developed to identify the different species based on the sugar assimilation profile and growth at different temperatures after 21 days. Main characteristics of S. brasiliensis, as described by Marimon et al. (2007), were that colonies grown on potato dextrose agar (PDA) at 30 °C attained a diameter of 15-38 mm after 21 days of incubation; sessile conidia were brown to dark brown and globose to subglobose; and the species was unable to assimilate sucrose and raffinose. Sporothrix globosa colonies grown on PDA at 30 °C attained a diameter of 18-40 mm after 21 days; sessile conidia were brown to dark brown and
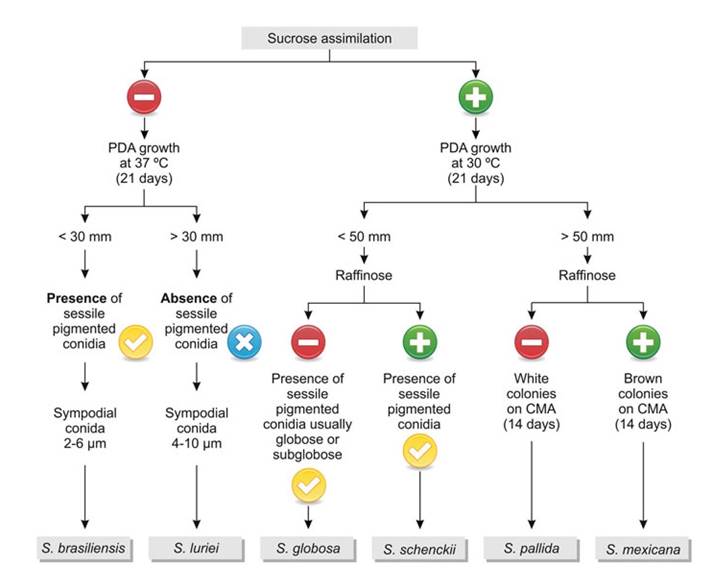
Fig. 10.3 Dichotomous key for identification of species of medical relevance in the genus Sporothrix. The criteria used for identification are sucrose and raffinose assimilation and colony size after 21 days of growth on potato dextrose agar (PDA), as well as micromorphological characteristics. CMA Corn Meal Agar
globose to subglobose; and the fungus was unable to assimilate raffinose, a key difference with S. schenckii. In turn, the S. mexicana colonies grown on PDA at 30 °C attained a diameter of 66-69 mm after 21 days; the sessile conidia were brown to dark brown and were predominantly globose, ovoidal, or ellipsoidal; and absence of assimilation of sucrose, raffinose, and ribitol. However, much phenotypic overlap has been reported in the literature (Oliveira et al. 2011; Rodrigues et al. 2013a; Camacho et al. 2015), and the dichotomous key should be used with caution to identify clinical Sporothrix species.
10.3