The Causative Agents of Feline Aspergillosis
In contrast to dogs with SNA, in which A. fumigatus sensu stricto causes over 95% of infections (Talbot et al. 2014), a more diverse range of Aspergillus species has been identified in FURTA (Table 15.1).
Overall, the Aspergillus viridinutans speciesTable 15.1 Causative agents in feline upper respiratory tract aspergillosis based on molecular identification
| Section | Species | Form of disease |
| Fumigati | A. fumigatus | SNA |
| A. lentulus | SNA | |
| A. fischeri | SOA | |
| A. thermomutatus | SNA, SOA | |
| A. felis | SNA, SOA | |
| A. parafelis | SOA | |
| A. udagawae | SOA | |
| A. wyomingensis | SOA | |
| Nigri | A. niger | SNA |
| Flavi | A. flavus | SNA |
SNA sinonasal aspergillosis, SOA sino-orbital aspergillosis
complex and the A. fumigatus species complex in section Fumigati are most frequently implicated in FURTA (Fig. 15.1).
Investigations into the causal agents of SOA in cats resulted in the recent discovery of a new species A. felis (Barrs et al. 2013), which is the most common cause of SOA followed by A. udagawae (Kano et al. 2008, 2013; Barrs et al. 2013, 2014, 2015). Two other molecular siblings of A. viridinutans, A. parafelis and A. wyomingensis, have been isolated from cats with SOA (Barrs et al.
2014; Talbot et al. 2017). The species A. viridinutans was reported to cause SOA in a cat from Japan (Kano et al. 2013); however the A. viridinutans sequences deposited on GenBank as HE578084 and AB24899 for which the isolate had greatest β-tubulin sequence homology with have been subsequently reclassified as A. felis on further phylogenetic analysis (Novakova et al. 2014). Other section Fumigati species isolated from cats with SOA include A. thermomutatus (Barrs et al. 2012) and A. fischeri (Kano et al. 2015).Aspergillus fumigatus s. str. and A. niger and relatives (section Nigri) are the most common isolates in SNA (Furrow and Groman 2009; Barrs et al. 2012, 2014, 2015; Whitney et al. 2005). Other section Fumigati species occasionally isolated from cats with SNA include A. lentulus, A. felis and A. thermomutatus (Barrs et al. 2013, 2014). Aspergillus flavus (section Flavi) was isolated from one cat with SNA in association with a plant foreign body (Barrs et al. 2015).
There are over 60 species of Aspergillus in section Fumigati with many new species additions in the last decade. That number is likely to continue to increase as molecular methods facilitate more accurate species delimitation, and since many more fungal species are thought to remain undiscovered. Cryptic species within section Fumigati cannot be reliably distinguished from A. fumigatus s. str. using phenotypic methods. However, accurate identification has important therapeutic implications since antifungal resistance profiles of cryptic species differ to that of A. fumigatus s. str., with some isolates having inherently higher MICs of azoles and other antifungal drugs. Molecular methods such as comparative sequence analysis and more recently MALDI-TOF are increasingly used for identification of human
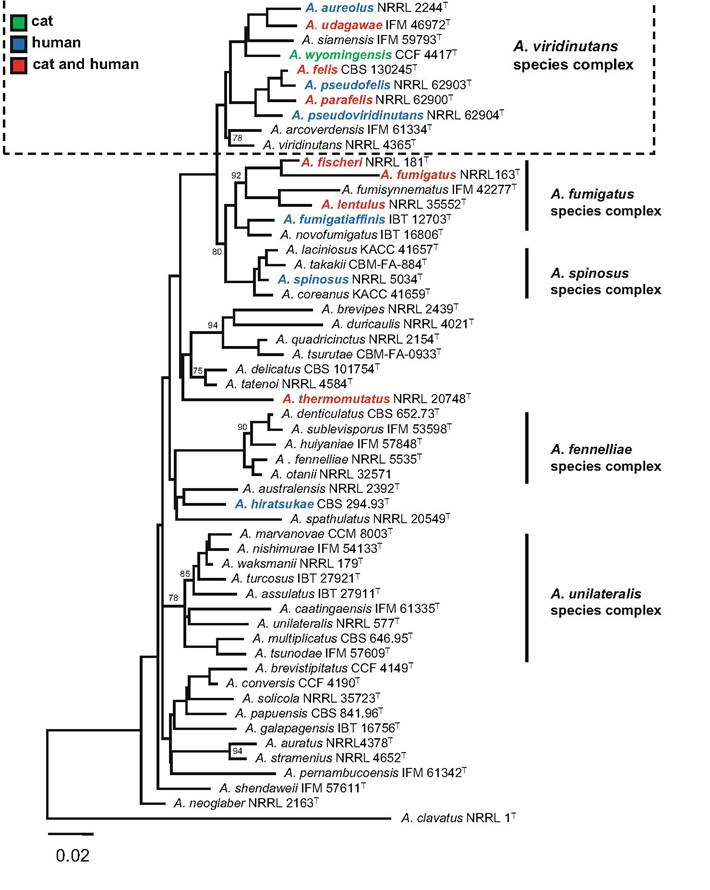
Fig. 15.1 Best-scoring maximum likelihood tree (kindly provided by Vit Hubka, Charles University, Prague) based on combined dataset of calmodulin and β-tubulin sequences showing the relationship of Aspergillus Viridinutans complex to other Aspergillus species from section Fumigati.
To filter both gaps and variable regions in the alignment, GBLOCKS v0.91b (Talavera and Castresana 2007) was used with less stringent selection allowing smaller final blocks and gap positions within the final blocks. The tree was constructed with the IQ-TREE version 1.4.0 (Nguyen et al. 2015). Dataset contained 58 taxa and a total of 908 characters of which 450 were variable and 292 parsimony-informative. Optimal partitioning scheme and substitution models were selected using PartitionFinder v1.1.0 (Lanfear et al. 2012) with setting allowing introns, exons and codonclinical Aspergillus isolates. Uptake of these methods in veterinary medicine has been slower. Misidentification of heterothallic cryptic species causing feline URTA including A. felis, A. udagawae and A. Ientulus is likely when only morphological typing methods are used (Barrs et al. 2012, 2013; Balajee et al. 2006). For example, in one case of SOA where A. fumigatus was reportedly identified using micro- and macromorphology, misidentification of a cryptic species was likely based on high MICs of amphotericin and itraconazole for this isolate (McLellan et al. 2006). Compared to A. fumigatus s. str., the MICs of amphotericin B for cryptic species including A. lentulus and A. udagawae are high (Balajee et al. 2006; Alcazar-Fuoli et al. 2008).
For molecular identification of section Fumigati species using comparative sequence analysis, a universal DNA barcode such as the nuclear ribosomal internal transcribed spacer (ITS) region is unreliable and should be combined with a secondary identification marker such as calmodulin, β-tubulin or the RNA polymerase II second largest subunit (RPB2) (Samson et al. 2014). Of these three, calmodulin is best for identification of clinical isolates since amplification of RPB2 is more difficult and amplification of β-tubulin can be complicated by the presence of paralogous genes and variable numbers of introns (Hubka and Kolarik 2012). For molecular identification of isolates from clinical samples, DNA can be extracted from fungal culture material for PCR and sequencing.
Alternatively, fungal DNA can be extracted directly from fresh or frozen clinical specimens or from formalin- fixed paraffin-embedded tissues (FFPET). However, DNA fragments from FFPET may be poor in quality and short, limiting the gene targets for amplification. The entire ribosomal DNA ITS1-5.8S-ITS2 region of approximately 600 nucleotides can easily be amplified from DNA extracted from fungal culture material, using primers ITS1 and ITS4 (Barrs et al. 2013). For FFPET, amplification of the shorter ITS1 (primers ITS1 and ITS2) region (~290 nt) or ITS2 (primers ITS3 and ITS4) region (~330 bp) is often possible (Barrs et al. 2012; Meason-Smith et al. 2017). In one study using amplification of the ITS2 region from FFPET for a range of fungal organisms, identification of fungi to genus level was possible in 65% of samples, and in 96% of these, the identified fungus was the same as that identified morphologically on histological examination (Meason-Smith et al. 2017).While all members of section Fumigati reproduce asexually by mitosis (anamorph), many are also capable of sexual reproduction by meiosis (teleomorph) under certain optimal growth conditions. Anamorphs are mould-like and are comprised of filamentous hyphae bearing mitotic spores. Teleomorphs comprise
Hoffman 2010; Barrs et al. 2012). There are surrounding zones of inflammatory cells including predominantly neutrophilic or eosinophilic infiltrates, activated and epithelioid macrophages and peripheral fibroblasts, lymphocytes and plasma cells. Inflammatory lesions can efface adjacent skeletal muscle and bone (Barrs et al. 2012). The globe is resistant to infiltration by fungal hyphae but may be affected by keratitis and anterior uveitis, and there may be invasion of adjacent structures including the nictitating membrane, eyelid, optic nerve and optic chiasm (McLellan et al. 2006; Giordano et al. 2010; Barachetti et al. 2009; Hamilton et al. 2000; Barrs et al. 2012). Brain involvement has been demonstrated on MRI in cats with advanced disease and seizures, but histological evaluation was not performed (Giordano et al.
2010; Smith and Hoffman 2010). Haematogenous dissemination is possible but has only been seen in one case by the author, a young Ragdoll cat that presented with DIA due to A. felis infection with sino-orbital, cervical lymph node and pulmonary involvement.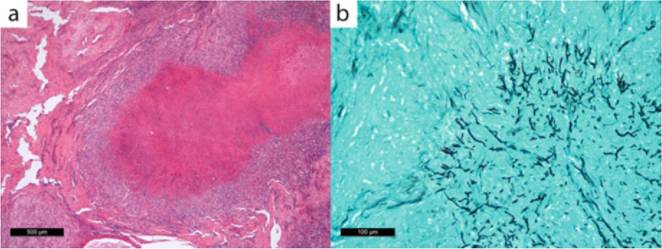
Fig. 15.3 Histopathology of an orbital fungal granuloma showing a central area of coagulative necrosis in which parallel-walled dichotomously branching, septate fungal hyphae are confined (a) and stain positively with Grocott's methenamine silver (b)
Aspergillus conidia that escape mucociliary clearance after inhalation are phagocytosed by macrophages and dendritic cells that express pattern recognition receptors (PRRs). PRRs recognise specific fungal epitopes or pathogen-associated molecular patterns (PAMPS) and activate Th1- Th2- and Th17-related intracellular molecular pathways triggering the production of proinflammatory and antiinflammatory cytokines, chemokines and their receptors and proangiogenic factors (Segal 2009). Single-nucleotide polymorphisms (SNPs) in PRRs in humans can increase susceptibility to IA, including SNPs in Toll-like receptors (TLRs) 1, 3, 4 and 6; in C-type lectin receptors (CLRs), Dectin-1 and DC-SIGN; in the secreted PPR mannan-binding lectin (MBL); and in PTX3 (Gresnigt et al. 2012; Romani 2011; Lupianez et al. 2016). In addition, SNPs in cytokines, chemokines and immune receptors increase susceptibility to IA (Lupianez et al. 2016). Whether similar genetic mutations could be associated with increased susceptibility to FURTA in brachycephalic cats of Persian lineage has not been investigated. Interestingly, Persian cats are also at increased risk for invasive dermatophyte infections (pseudomycetomas) (Miller 2010). Expression of TLR 2, 4 and 9 mRNA was upregulated in the nasal mucosa of dogs with SNA, prompting a mutation analysis of these genes. However, SNPs correlating with an increased susceptibility to canine SNA were not identified (Mercier et al.
2012, 2014).Conformational abnormalities of the upper respiratory system may contribute to the increased risk of URTA in brachycephalic breeds of cats. Congenital shortening of the facial and neurocranial bones, together with dorsal rotation of the jaw, results in deformation and displacement of ventral nasal and ethmoid turbinates as well as impaired nasolacrimal drainage (Schlueter et al. 2009). Decreased sinus aeration and respiratory secretion drainage secondary to infection, polyps and allergic rhinosinusitis are a risk factor for invasive FRS in humans (Siddiqui et al. 2004). Similarly, impaired drainage of nasal secretions has been proposed as a risk factor for fungal colonisation in brachycephalic cats (Tomsa et al. 2003). By contrast, in dogs with SNA, brachycephalic dogs are under-represented, and disease is most frequent in dolichocephalic and mesaticephalic breeds (Peeters and Clercx 2007). Additional risk factors such as previous viral respiratory infection or recurrent antimicrobial therapy may increase the risk of sinonasal fungal colonisation in brachycephalic cats (Tomsa et al. 2003; Barrs et al. 2012; Goodall et al. 1984).
A role for species-specific fungal virulence factors in the development of invasive FURTA is suspected since different fungal species are implicated in SNA and SOA in cats (Barrs et al. 2013). This has been demonstrated to some extent in vitro. Galleria mellonella larvae and two murine hosts (BALB/c mice immunosuppressed with hydrocortisone and mice with chronic granulomatous disease (GCD)) were inoculated with conidial suspensions of A. fumigatus and A.felis (Sugui et al. 2014). The A. felis type strain (isolated from a cat with SOA) showed significantly higher virulence in G. mellonella larvae than A. fumigatus. The species A. felis was slightly more virulent in immunosuppressed BALB/c mice than A. fumigatus but was less virulent in mice with GCD. Aspergillus fumigatus caused 100% mortality in GCD mice in 11 days, whereas A. felis caused no fatalities, demonstrating decreased virulence in hosts deficient in production of reactive oxygen species.
Secondary fungal metabolites or extrolites are numerous, often bioactive and species-specific and may contribute to infection and invasion. For example, gliotoxin inhibits immune responses, phagocytosis and angiogenesis and is produced by A. fumigatus and A. thermomutatus. Fumagillin also suppresses immune responses, neutrophil function and angiogenesis and is synthesised by A. felis, A. fumigatus and A. udagawae. Sulochrin, an eosinophil activation inhibitor, is synthesised by these same three species, as well as by A. lentulus. Helvolic acid can adversely affect respiratory epithelium and macrophage metabolism and is synthesised by many of the section Fumigati species that cause FURTA (Frisvad and Larsen 2016). The role of many extrolites in fungal pathogenicity remains to be elucidated.
15.6