The Etiological Agent
LCD was diagnosed as a systemic infection associated to black yeasts, supporting preliminary identification of the causative agent as a species of Exophiala (Boeger et al. 2005).
The hypothesis of a viral origin, proposed by Schaeffer-Novelli et al. (2004), was refuted due to the lack of evidence from histological analyses, by molecular diagnostic tests for a number of known shrimp viruses (white spot syndrome virus [WSSV], infectious hypodermal hematopoietic necrosis virus [IHHNV], hematopancreatic parso-like virus [HPV], mesodermal baculovirus [MBV], baculovirus penaei [BP], Taura syndrome virus [TSV], gill-associated virus [GAV]), and by experiments of artificial infection which failed to detect any evidence of viral infection (Orelis-Ribeiro et al. unpublished).Putative causative agents of LCD in the mangrove coast of Brazil were isolated from direct culturing of tissues (hepatopancreas and heart) from moribund mangrove crabs (U. cordatus) showing signs of the disease (Boeger et al. 2007). A black yeastlike species was isolated from crabs in all stages of the disease. The agent was described as a new species, Exophiala cancerae (de Hoog et al. 2011) (Fig. 11.4). In two cases, a coinfection by a second melanized, fonsecaea-like fungus was detected. Sequences of the ribosomal operon were obtained for isolates originating from
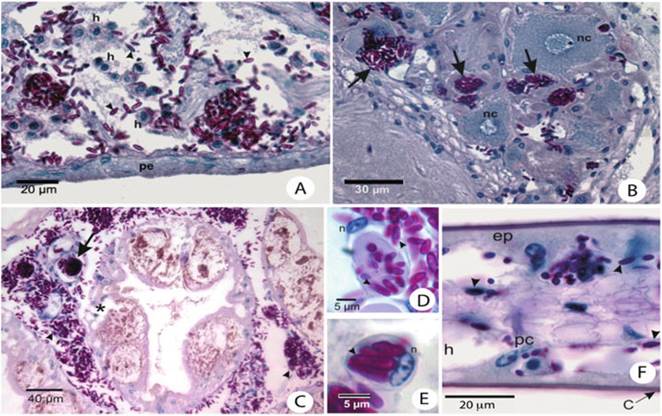
Fig. 11.3 Histopathology of Ucides cordatus infected showing signs of lethargic crab disease: (a) Heart of a moribund mangrove-land crab with LCD. Disruption and disorganization of cardiacmuscle fibers associated with the yeast-like cells (arrowheads) with extensive hemocytic infiltration (h). Note that pericardia (pe) are free of yeast-like cells. PAS and HE stains. (b) Neurosecretory area of the thoracic ganglion of a moribund mangrove-land crab with LCD with hemal sinuses congested by yeast-like cells (arrows).
nc neurosecretory cells. PAS and HE stains. (c) Large hemocytic agglutination congesting the hemal sinus between tubules of the hepatopancreas, with both yeast cells (asterisk) and hyphae (arrows). PAS and HE stains. Yeast-like cells (arrowhead) and hemocytic encapsulations (arrow) occlude hemal sinuses of the hepatopancreas. The hepatopancreas caecum shows signs of necrosis (asterisk). PAS and HE stains. (d) RI cell with numerous yeast-like cells (arrowheads) in the connective tissue associated with the exoskeleton. n nucleus of the cell. Periodic acid Schiff (PAS) and HE stain. (e) Phagocytic cell (probably a hyalinocyte) of the connective tissue associated with the exoskeleton with numerous yeast-like cells (arrowhead). n nucleus of the cell. PAS and HE stains. (f) Gills of mangrove-land crab with LCD in intermediate stage of infection. Although many yeast-like cells (arrowheads) are visible, most of its components are relatively intact. c cuticle, pc pillar cell, ep epithelium, h hemal lacuna. PAS and HE stainsdifferent crab tissues, and also this species appeared to represent an unknown taxon, which was described as Fonsecaea brasiliensis by Vicente et al. (2012) (Fig. 11.4).
LCD was associated with the presence of a black yeast-like fungus, E. cancerae, a member of the ascomycete order Chaetothyriales comprising the black yeasts and their filamentous relatives, as was proven studies using light and electron microscopy, behavioral and experimental tests, and molecular phylogenetics. Species of this order are regularly encountered as causative agents of cutaneous and systemic disorders in humans and cold-blooded animals (Nyaoke et al. 2009; de Hoog et al. 2011; Kano et al. 2000). Human infections range from commensalism or mild cutaneous infection to fatal neurotropism or osteotropism, sometimes with extracutaneous dissemination with severe mutilation. Waterborne vertebrates such as toads, frogs, and turtles are particularly susceptible to infection by Exophiala or fonsecaea-like species (de Hoog et al.
2011). Some species are host-specific to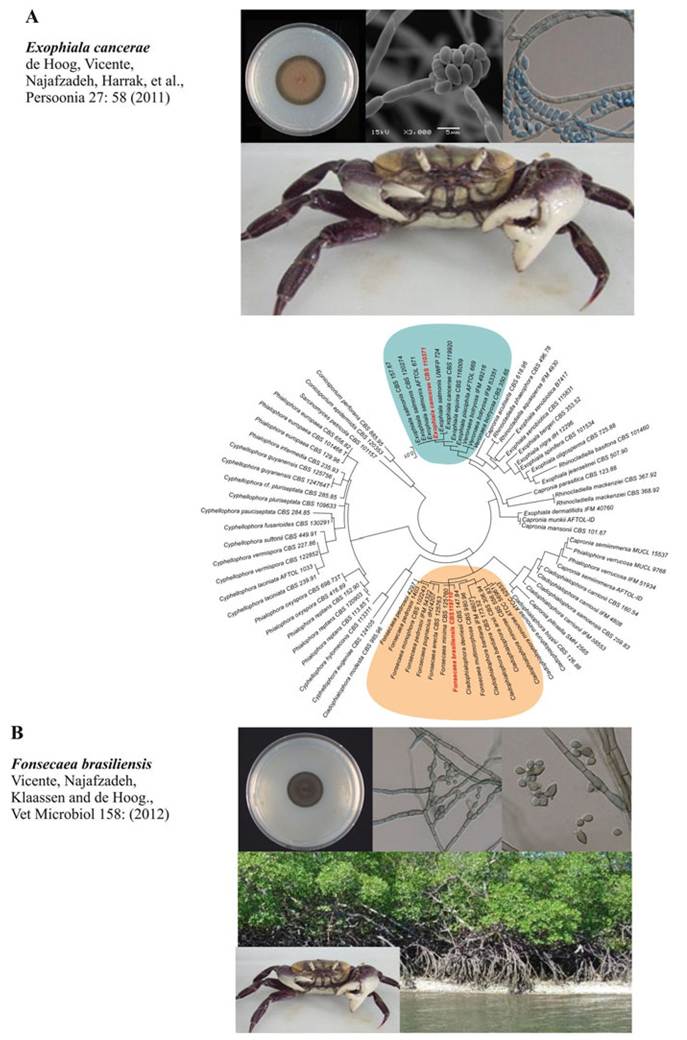
Fig. 11.4 Phylogenetic tree based on confidently aligned of LSU sequences constructed with maximum likelihood implemented in RAxML. Colored rectangles indicate main clades recognized within by Vicente et al. (2012) and de Hoog et al. (2011) which grouped the two agents of lethargic
particular fish taxa (Nyaoke et al. 2009). Virulence factors are largely unknown, but the ability to assimilate alkylbenzenes that are present in sweat and in nervous tissues of mammals as well as in the toxic skin of amphibians, has been suggested degradation of monoaromates is a rare property in the fungal Kingdom (Prenafeta-Boldu et al. 2006; de Hoog et al. 2011). Older literature data demonstrate that black yeast infections taking epizootic dimensions are relatively common in fish and frogs (e.g. Fijan 1969; Richards et al. 1978; Reuter et al. 2003). Several opportunistic species are involved (e.g. Manharth et al. 2005; Nyaoke et al. 2009; de Hoog et al. 2011) which are morphologically very similar. Invertebrate hosts in moist habitats may also be infected. In addition to crabs, Exophiala species have been reported from earthworms (Vakili 1993) and from mussels (Dover et al. 2007).
Fonsecaea brasiliensis and E. cancerae are members of rather distant clades within the Chaetothyriales, as proven by phylogenetic analysis of partial large subunit (LSU) rDNA data (Fig. 11.3). Isolates of E. cancerae belong to a clade of waterborne species of Exophiala, which are potentially able to cause infectious diseases in cold-blooded animals (de Hoog et al. 2011). Fonsecaea brasiliensis from crab clustered with Cladophialophora devriesii and C. bantiana. The former species was originally described from a fatal disseminated infection in a healthy appearing Caribbean woman (Gonzalez et al. 1984), although an inherited immune disorder such as a CARD9 defect cannot be excluded, while C.
bantiana is an important agent with neurotropic invasion in human (de Hoog et al. 2009).A multilocus analysis based on sequences of ITS (rDNA internal transcribed spacer) and partial sequences of translation elongation factor 1-a (TEF1), β-tubulin (BT2), as well as fingerprint profiles generated by amplification of restriction length polymorphism (AFLP) were used to confirm the identity of F. brasiliensis from crab and its relationship to C. devriesii (Vicente et al. 2012).
Exophiala cancerae was present in all crabs exhibiting clinical signs of LCD investigated and is considered to be the main agent of the disease. The natural niches of both species remain unclear. Fonsecaea brasiliensis was encountered coinfecting mangrove crab, but Vicente et al. (2012) could not establish whether severity of the disease increased with double infections. Environmental isolation studies showed that F. brasiliensis was abundantly present in the environmental niches known as crab habitats but also in plant debris in the Brazilian rainforest (Vicente et al. 2012, 2014; Guerra et al. 2013) suggesting that F. brasiliensis was a general opportunist of moribund crabs. In contrast, E. cancerae could not be found in the local environment (Vicente et al. 2012), not even from focus areas of the disease, suggesting a certain degree of host specificity. Remarkably, however the species was found elsewhere in water, once causing an outbreak in toads (J. Cunningham, pers. comm.), and was also repeatedly isolated from human nails (de Hoog et al. 2011).
Fig. 11.4 (continued) crab disease (LCD), Fonsecaea brasiliensis and Exophiala cancaerae showed in red letter in the tree. The macro/micromorphology and ecological aspects of the two agents of LCD are presented: (A) colony, conidiophores apparatus, and crab host; (B) colony, conidiophores apparatus, and environmental niche/crab host
Experimental infections of E. cancerae (Orelis-Ribeiro et al. 2011) and of F. brasiliensis (Vicente et al.
2012) provided strong evidence that the former species is likely the major causative agent of LCD (Fig. 11.5). When infected with E. cancerae, crab dying is beginning to be observed between the 7th and the 10th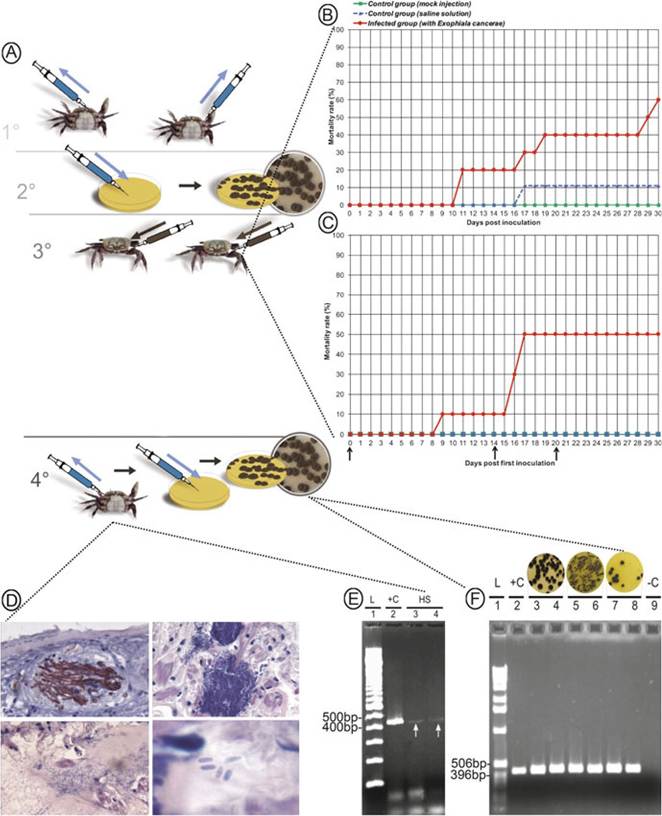
Fig. 11.5 Diagram illustrating the fulfillment of Koch’s postulates on the context of LCD. (A) Koch’s postulates; (B and C) Mortality curves for crabs that were subjected to a single and multiple experimental infection;(D) Histology of experimentally infected crabs; (E) Molecular diagnosis of infected tissues; (F) Molecular identification of colonies recovered from the tissue of artificially infected crabs. Adapted from Orelis-Ribeiro et al. (2011)
day post infection and reaches up to 60% mortality between days 17 and 33, when the experiment was terminated. The diseased crabs, inoculated with E. cancerae, reproduced the clinical signs of LCD observed in the mangroves. None or very limited mortalities were observed in the control groups (mock or saline-solution injections). Mortality of crabs injected with a conidial suspension of F. brasiliensis was significantly less pronounced and began 27 days after infection, reaching 40% by the end of the experiment. Two weeks after plating the collected tissues from crabs that died during the experiment on Mycosel medium, resulting colonies were identified down to genus level by morphology and to species level by the use of specific PCR primers. The lower virulence of F. brasiliensis supports its nature as a secondary invader in natural infections, probably taking advantage of weakened crabs initially infected by E. cancerae.
11.4