Action Potentials Begin at the Axon's Initial Segment and Spread Down the Entire Length of the Axon
Both EPSPs and IPSPs on the postsynaptic membrane are ultimately the result of an action potential and synaptic transmission from a presynaptic cell. However, these postsynaptic potentials decrease in magnitude as they spread along the postsynaptic cell membrane.
Because many neurons and muscle cells are long, the cell needs a mechanism for sending an electrical signal from its information-receiving end on the postsynaptic dendritic and soma membrane to the information-transmitting zone at the terminals of the often- Iengthy axon. This is accomplished by an explosive event called an action potential, a regenerative electrical signal that begins at the axon’s initial segment, results from competing EPSP and IPSP membrane potential changes, and spreads down the length of the axon without decreasing in magnitude.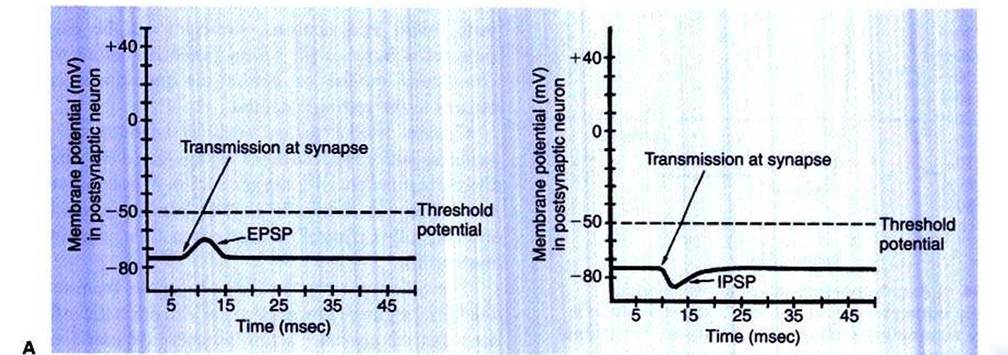
FIGURE 4-3 Postsynaptic potentials. Ar Excitatory postsynaptic potential (EPSP) drives the membrane potential toward threshold. Br Inhibitory postsynaptic potential (IPSP) drives the membrane potential away from threshold.
B
EPSPs and IPSPs can respectively summate on the postsynaptic membrane to produce larger changes in membrane potential than either signal alone. At the axon’s initial segment, the arriving EPSPs and IPSPs are integrated. If only a few EPSPs arrive at the axon’s initial segment, its membrane potential is not made sufficiently positive to reach its threshold potential (often 10-20 mV more positive than the resting potential) for triggering an action potential. However, if many more EPSPs than IPSPs arrive, the initial segment’s membrane potential is made positive enough to reach its threshold potential, and an action potential is created. This action potential is the result of the sequential opening of voltage-gated membrane channels first to sodium and shortly thereafter to potassium.
The action potential is characterized by explosive changes in the membrane potential when this value reaches the threshold potential: First, a dramatic and swift depolarization of the membrane potential occurs, in which the inside of the cell actually becomes more positively charged than the outside, followed by a repolarization of the membrane, in which the membrane potential falls back toward the resting potential. The depolarization phase of the action potential is caused by the immediate and extensive opening of voltage-gated Nat channels and the consequent influx of Na4 ions as they attempt to flow toward their equilibrium. As the action potential’s
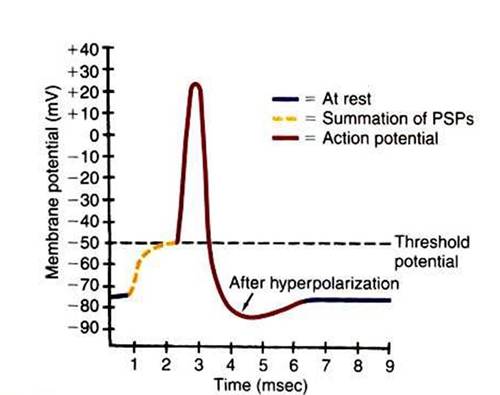
FIGURE 4-4 ■ Axon's membrane potential changes dramatically during an action potential. After threshold is reached by summating postsynaptic potentials, the axonal membrane depolarizes, repolarizes, hyperpolarizes, and then returns to its original resting potential. (Modified from Sherwood L: Human physiology: from cells to systems, St Paul, 1989, Wadsworth.)
depolarization phase continues, the voltage-gated Na+ channels are spontaneously inactivated, and the voltage-gated K+ channels, which open with a longer delay than the Na’ channels, begin to allow even more K* ions to exit as they move closer to their equilibrium state. This brings depolarization to a halt and allows repolarization to occur. As repolarization continues, the membrane potential moves temporarily beyond its resting level to a hyperpolarized state. This hyperpolarization is attributable to the flow of K’ ions out through the voltage-gated Kh channels, in addition to the flow out through the K' leak channels, bringing the membrane potential even closer to the K, equilibrium potential (-90 mV) than at rest. The membrane potential eventually returns to its resting state as the K* voltage-gated channels gradually close.
The whole action potential takes about 2 to 3 msec in many neurons but longer in many muscle cells. Figure 4-4 illustrates this sequence of events in a neuron.An analogy may be helpful for understanding these difficult concepts. Imagine the resting neural membrane as a toilet. The toilet has stored potential energy by filling its water tank. (The neuron has done so by generating the resting membrane potential.) If the handle of the toilet is pushed down only briefly, for a short distance, some water runs into the toilet, but the flush cycle is not initiated. (This is similar to an EPSP without the action potential.) However, if the handle is pushed down far enough and held down long enough, a critical threshold is reached, the flush cycle is triggered, and it must run its course, including the refilling of the tank, before another flush cycle can be started. The action potential is analogous to this flush cycle. It is triggered once a critical depolarization threshold is reached. It usually must run its course, including reestablishing the resting membrane potential, before another action potential can be initiated. Because the flush cycle takes a finite amount of time, only a limited number of flush cycles could be completed in an hour, even if the toilet were flushed again each time the tank refilled. Similarly, because the action potential also has a finite duration, there is a limit to the number of action potentials per second that can be generated on an axon. (However, for both toilets and neurons, strategies can be employed to produce a flush or an action potential before the tank is completely refilled or before the membrane completely returns to the resting potential.)
Certain animal toxins, such as Ietrodotoxin from the Japanese puffer fish, can block voltage-gated Na, channels and therefore interfere with the generation of action potentials on axons. Many local anesthetics (e.g., lidocaine), which are used in a controlled, clinically efficacious manner, work by a similar mechanism.
The action potential actively propagates from its origin at the initial segment down the axon. The dramatic influx of Na+ ions that accompanies action potential depolarization of the initial segment’s membrane results in the passive spread of these positive charges toward the adjacent resting segment of membrane. This migration of positive charge on the inner
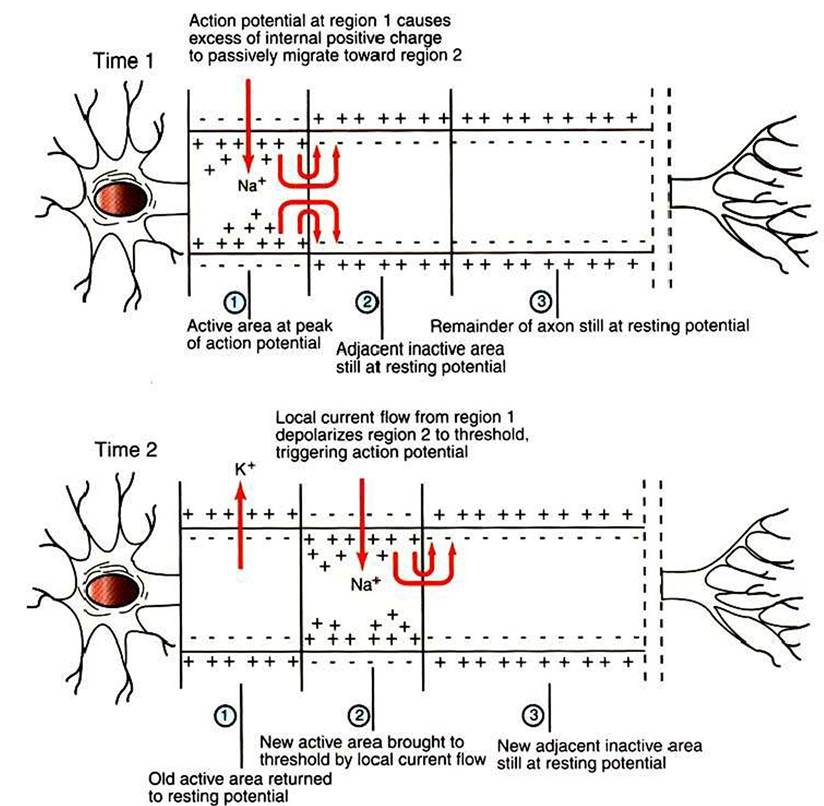
FIGURE 4-5 Action potential, first generated in the axon's initial segment (Time 1, region 1), moves down the unmyelinated axon as positive charges passively migrate to the immediately adjacent membrane to trigger an action potential there (Time 2, region 2). (Redrawn from Sherwood L: Human physiology: from cells to systems, St Paul, 1989, Wadsworth.)
surface of the membrane, called an electrotonic current, depolarizes this adjacent segment to threshold, causing voltage-gated Na4 channels to open. This causes an action potential to develop there, which triggers a similar cycle in its adjacent membrane, and so on down the axon. In this way an action potential spreads from the axon’s initial segment down to the presynaptic terminal at the axon’s far end (Figure 4-5).
The speed with which the action potential is conducted down the axon varies. The internal diameter and the degree of myelination of an axon play a critical role in determining this action-potential conduction velocity. In a small-diameter, unmyelinated axon, the conduction velocity is relatively slow (e.g., 0.5 meter/second); conduction velocities of greater than 90 m/sec (so that a distance as long as a football field is traveled in 1 second), however, are known to occur in large- diameter, heavily myelinated axons. This occurs because the passive electrotonic current, responsible for triggering the action potential at the next adjacent patch of axonal membrane, travels faster and farther along wider axons or along myelinated patches of axon.
In myelinated axons, exchange of ions across the membrane, and thus generation of the action potential, can only occur at the bare nodes of Ranvier, where a high density of voltage-gated Na, channels is found. Given the rapid spread of electrotonic current along the myelinated patches (internodes) and the comparatively slow process of ion exchange at the nodes, the action potential functionally jumps from node to node (saltatory conduction) in myelinated axons (Figure 4-6).The normal facilitation of action-potential conduction velocity by myelin can be appreciated by considering diseases that attack myelin, such as acute idiopathic polyradiculoneuritis (ucoonhound paralysis”). Slowing of evoked electrical signals along sensory and motor nerves and depressed spinal reflexes are associated with this condition.
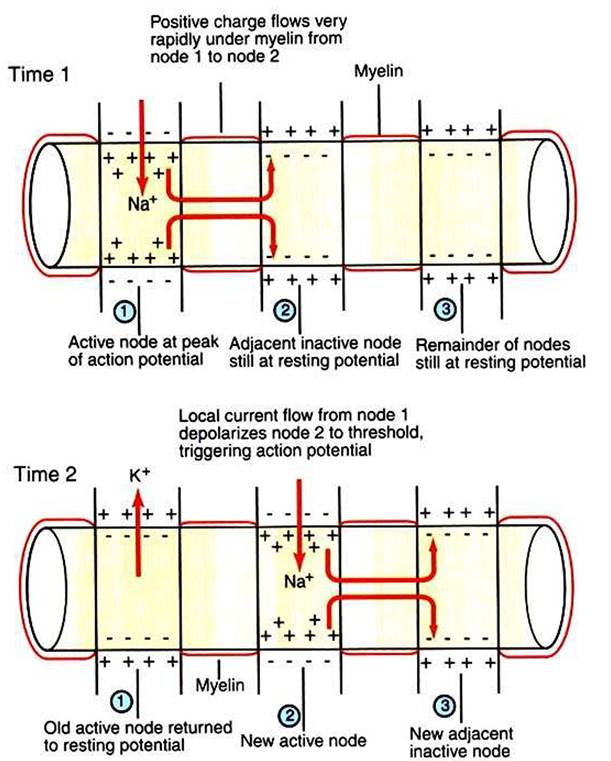
FIGURE 4-6 Saltatory conduction of action potentials in myelinated axons is faster than action-potential conduction in unmyelinated axons because the passive local current flows very rapidly under the myelin to trigger an action potential at the next node. Thus the action potential seems to jump functionally from node to node. (Modified from Sherwood L: Human physiology: from cells to systems, St PauL 1989, Wadsworth.)