Anaerobic Conditions in the Rumen Result in Metabolic Activities Leading to the Production of Volatile Fatty Acids
When carbohydrate material enters the rumen or colon, it is attacked by hydrolytic microbial enzymes, in the case of insoluble carbohydrates, attack requires the physical attachment of bacteria to the surface of the plant particle, with the enzymes themselves part of the surface coating of the bacteria.
Enzymatic action liberates glucose, other monosaccharides, and short-chain polysaccharides into the fluid phase, outside the microbial cell bodies. Although free in solution, these products of microbial enzyme action do not become immediately available to the host animal; rather, they are quickly subjected to further metabolism by the microbial mass. Glucose and other sugars are absorbed into the cell bodies of the microbes.Once within the microbial cells, glucose enters the glycolytic, or Embden-Meyerhof, pathway. This is the same glycolytic pathway that exists in mammalian cells, and as in mammalian tissues, catabolism of glucose through this pathway yields two molecules of pyruvate Ioreach Inoleculeofglucose metabolized. In the process, two molecules of oxidized nicotinamide adenine dinucleotide (NAD) are reduced to NAD hydrogen (NADH), and two molecules of adenosine triphosphate (ATP) are formed from adenosine diphosphate (ADP). The potential energy represented by the z∖TP formed in this reaction is not directly available to the host animal but is the major source of energy for maintenance and growth of microbes.
If fermentative digestion were to occur under aerobic conditions, which it does not, the pyruvate produced by the glycolytic process would enter the citric acid (Krebs) cycle and would be metabolized to carbon dioxide and water, as occurs under the aerobic conditions in mammalian cells. Furthermore, in an aerobic system, the NADH produced would be oxidized in the cytochrome oxidase system with additional production of ATP and the regeneration of NAD.
However, fermentative digestion is not an aerobic system; on the contrary, it proceeds in a reductive, highly anaerobic environment. Therefore a different mechanism must be provided for the oxidation of NADH and other reduced cofactors. If such a mechanism were not available, all the oxidized cofactors present would soon be reduced, and metabolism would come to a halt. Because no atmospheric oxygen is available, some other compound must serve as an “electron sink" for the oxidation of enzyme cofactors.In fermentative digestion, pyruvate can act as an electron sink, being further reduced to provide for regeneration of NAD and the general removal of excess electrons, with an additional yield ot ATP. Also, carbon dioxide can be reduced to methane, accepting electrons for the regeneration of NAD. Figure 31-1 illustrates the metabolic pathways of these reactions. These pathways lead to the major end products of the fermentative digestion of carbohydrate, the volatile fatty acids (VFAs). The primary VFAs are acetic acid, propionic acid, and butyric acid; the VFAs are often referred to as their dissociated ions: acetate, propionate, and butyrate, respectively. Other quantitatively minor but metabolically important
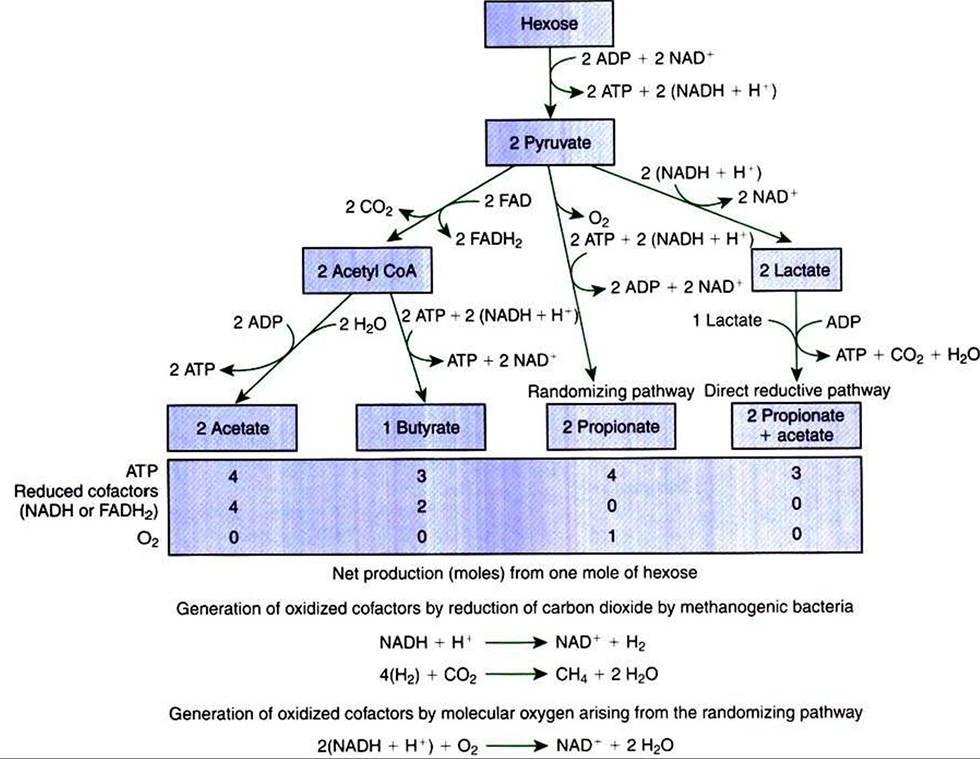
FIGURE 31-1 Pathways of volatile fatty-acid (VFA) production by the rumen or colonic biomass.The production of methane is necessary for the production of oxidized cofactors in the pathways leading to acetate and butyrate production, but not in the pathways leading to propionate production. The production of oxygen by the randomizing pathway results in the net production of oxidized cofactors. ADPAdenosine diphosphate; ATP, adenosine triphosphate; NAD, nicotinamide adenine dinucleotide; FAD, flavin adenine dinucleotide; H, hydrogen; CoA, coenzyme A; CO2. carbon dioxide.
VFAs are valeric acid, isovaleric acid, isobutyric acid, and 2-methylbutyric acid.
Figure 31-2 shows the chemical structures of the VFAs.Production of propionic acid from pyruvate results in the efficient regeneration of NAD with no net production of NADH. In fact, production of available oxygen by the randomizing branch of the propionic acid pathway leads to oxidation of excess NADH originating from the acetic or butyric acid pathways (see Figure 31-1). The production of acetic acid leads to the efficient generation of ATP but, in contrast to the production of propionic acid, does not result in the regeneration of NAD from NADH. In the acetic acid pathway, excess NADH is produced. In this case, NAD is regenerated by the formation of free hydrogen, which is subsequently used to reduce carbon dioxide to methane and water (see Figure 3∖-∖i lower portion).
Thus a direct relationship exists between acetic acid production and methane production; as the amount of pyruvate entering the acetic acid pathway increases, there must be a concomitant rise in methane production. Likewise, a reciprocal relationship exists between methane production and propionic acid production; as pyruvate is diverted to propionic acid production, there is less need for methane synthesis. These relationships are shown in the stoichiometric equations of Box 31-2. These reactions do not, however, fully describe the flow of hydrogen, or reducing substances, in rumen or colonic metabolism. The chemical reactions of fermentation are extremely complex and interdependent, and NADH can donate its electrons to reactions other than those described in Box 31-2, such as the synthesis of microbial protein and the saturation of unsaturated fatty acids.
In the rumen, methane production is facilitated by methanogenic bacteria, such as Methanobacterium ruminantium. This fragile bacterium is sensitive to changing conditions in the rumen. When conditions are unfavorable for the survival of M. runιinantiunιi methane production is reduced, shifting the metabolic pathways toward propionic acid production.
Some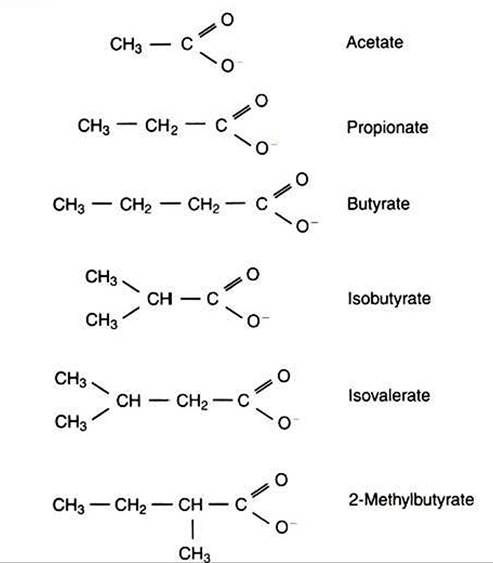
FIGURE 31-2 ■ Chemical structures of the major volatile fatty acids (VFAs) produced by fermentative digestion.
conditions that suppress methanogenic species are high levels of feed intake, use of finely ground or pelleted feeds, and high- grain or high-starch diets. Under these circumstances the rate of methane production is reduced, resulting in a lower rate of acetic acid production with a concomitant increase in the propionic acid production rate.
The proportional rates at which acetic acid, propionic acid, and butyric acid are produced are reflected in their relative concentrations in the rumen fluid. The relative concentrations of the VFAs have important nutritional and metabolic consequences, and although seldom measured for medical purposes, VFA concentrations are frequently reported in research literature. Typically, the ruminal acetic/propionic/butyric acid
Box 31-2
Theoretical Stoichiometric Carbon-Hydrogen Balance Equations Describing Conversion of Glucose in Rumen

From Van Soest PJ: Nutritional ecology of the ruminant, Ithaca, NY, 1982, Cornell University Press.
*Note that in Case 1, the acetate/propionate ratio is 1:1 and the methane/glucose ratio 1:3, whereas in Case 2, the acetate/propionate ratio is 3:1 and the methane/glucose ratio 3:5.
concentration ratio in ruminants ranges from 70:20:10 for animals eating high-forage diets to 60:30:10 for animals eating high-grain diets. One must remember that these values represent relative proportions and not absolute amounts. The total amount of VFA produced with a high-starch diet is usually much higher than that produced with a high-fiber diet, such that total acetic acid production may be higher with a high-starch diet than with a high-fiber diet, even though the acetic acid production relative to the other VFAs may be reduced. Figure 31-3 illustrates this principle.
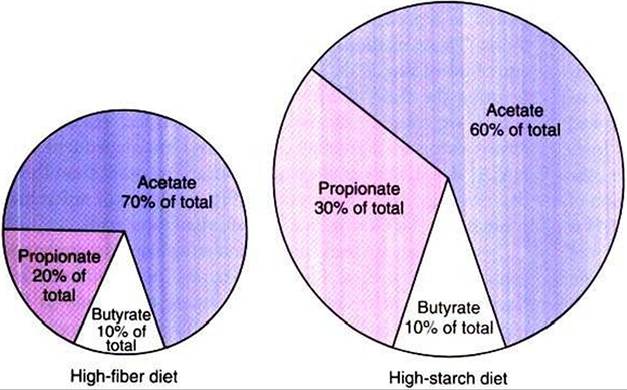
FIGURE 31-3 VFA production on high-fiber and high-starch diets. Although the percentage of acetate is lower on the high-starch diet, the total amount of acetate produced is greater on the high-starch diet. In contrast, propionate increases in both amount and proportion on the high-starch diet.