BASIC PLAN AND DEVELOPMENT
The trunk is the large part of the carcass that remains after the removal of the head and neck, the tail, and the forelimbs and hindlimbs; in common speech, it is the body of the animal (Figure 2-1).
It consists of three segments—thorax, abdomen, and pelvis—which are not clearly divided externally. Each is bounded by the body wall, and each contains a cavity, or a potential cavity, since, in life, the space is more or less obliterated by the close apposition of the walls and contents. The thoracic cavity lies cranial to the diaphragm, a domed sheet of muscle and tendon with a peripheral attachment to the body wall and a free center that bulges cranially. The abdominal cavity lies caudal to the diaphragm and corresponds to the belly. It communicates freely with the pelvic cavity within the enclosure of the bony pelvis (Figure 2-2).The dorsal part of the body wall that roofs the thoracic, abdominal, and pelvic cavities is known as the back. It is formed by the vertebral column and associated muscles, which are structures that also extend through the neck and tail. It is therefore convenient, if not entirely appropriate, to consider the vertebrae and associated structures of the neck and tail in this section. The structures of the ventral part of the neck are included with the head.
The neck, back, and tail exhibit a serial repetition of like elements, most notably the vertebrae. This apparent segmentation is, as reference to a young embryo shows (Figure 2-3), a legacy of the somites, the blocks into which the paraxial mesoderm is segregated to each side of the neural tube and notochord. The appearance in the adult is somewhat misleading; the vertebrae are, in fact, each formed by contributions from two somites of each side and are therefore more accurately described as intersegmental. Together with the ribs and sternum, they are produced from the medial portions of the somites known as sclerotomes.
The muscles of the vertebral column are derived from the lateral portions of the somites, the myotomes. Many adult muscles are polysegmental and combine contributions from several or even many myotomes, but certain groups of deeper units retain the unisegmental pattern. Because the vertebrae are intersegmental, even the shortest muscles bridge, and thus can move, the joint between two successive bones.Early on, each myotome attracts a single nerve (Figure 2-3/8) that grows out from the adjacent neural tube; from this, it follows that the motor innervation of the muscles is also segmental and that polysegmental muscles will have a multiple innervation. A similar pattern is apparent in the sensory innervation of the skin. It was formerly believed that the connective tissue component of the skin, the dermis, derived exclusively from third portions of the somites, the dermatomes. Cells from these were supposed to migrate to underlie specific regions of the surface ectoderm. This ordered pattern of migration is now in question, and it is thought that the dermis may be, in part, produced through mesenchyme differentiating in situ. Be that as it may, a segmental innervation of skin (Figure 2-4) exists in the
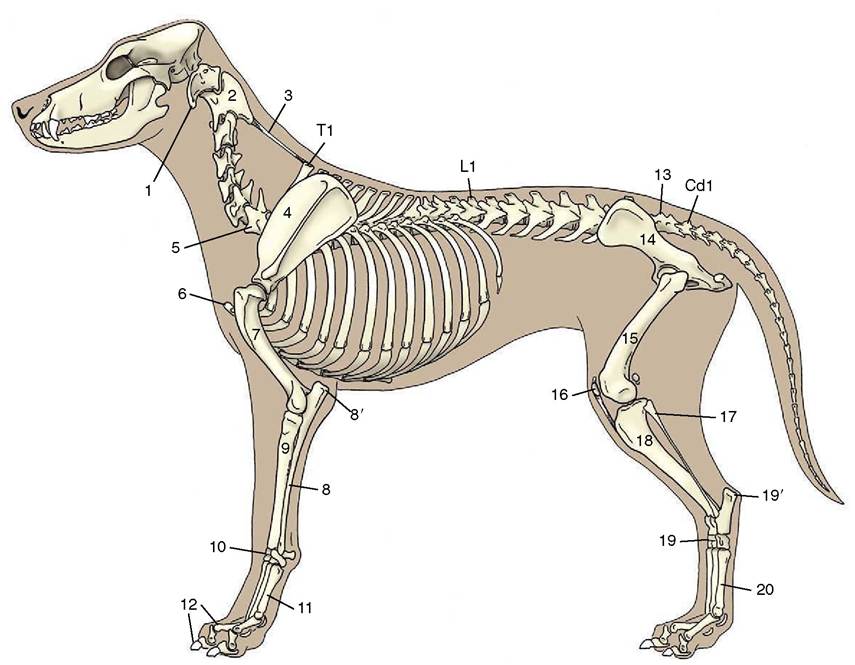
Figure 2-1 The skeleton of the dog. 1, Wing of atlas, first cervical vertebra (C1); 2, spine of axis (C2); 3, ligamentum nuchae; 4, scapula; 5, last cervical vertebra (C7); 6, cranial end (manubrium) of sternum; 7, humerus; 8, ulna; 8', olecranon (point of elbow); 9, radius; 10, carpal bones; 11, metacarpal bones; 12, proximal, middle, and distal phalanges; 13, sacrum; 14, hip bone (os coxae); 15, femur; 16, patella; 17, fibula; 18, tibia; 19, tarsal bones; 19', calcanean tuber (point of hock); 20, metatarsal bones; T1, L1, and Cd1, first thoracic, lumbar, and caudal (tail) vertebrae.
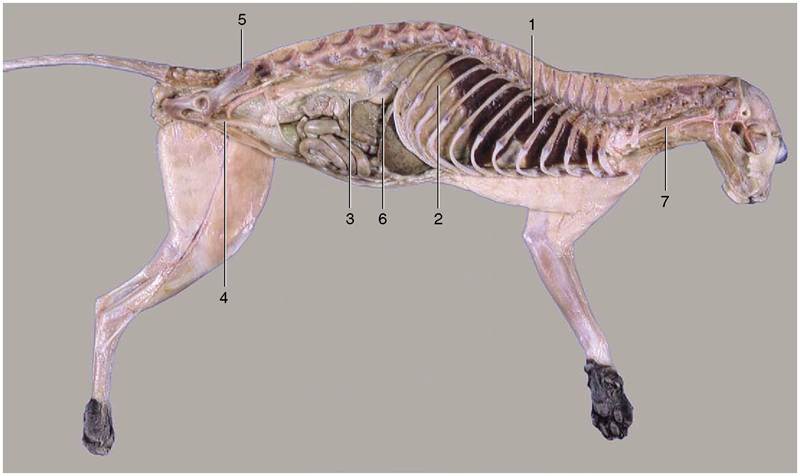
Figure 2-2 The thoracic, abdominal, and pelvic cavities of a cat; viewed from the left.
1, Thoracic cavity (with lung); 2, diaphragm; 3, abdominal cavity; 4, pelvic cavity; 5, sacrum; 6, right kidney; 7, esophagus.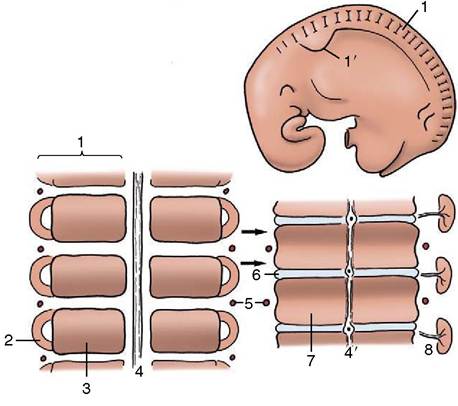
Figure 2-3 Segmentation of the paraxial mesoderm shown in a 10-mm bovine embryo (above) together with two stages in the development of the vertebrae and related vessels and nerves. The arrows show the formation of each vertebra from two pairs of adjacent somites. 1, Somite; 1', forelimb bud; 2, myotome; 3, sclerotome; 4, notochord; 4', notochord giving rise to the nucleus pulposus in the center of the intervertebral disk (6); 5, intersegmental artery; 6, intervertebral disk; 7, body of vertebra; 8, myotome with segmental nerve.
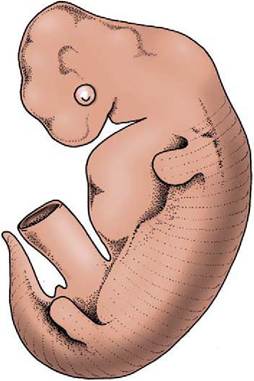
Figure 2-4 Embryo with "dermatomes" indicating the segmental innervation of the skin.
adult that is very regular in some places and less so in others. The bands of skin that are the provinces of particular pairs of spinal nerves are also known as dermatomes. Many overlap their neighbors. The associations between these bands and particular sensory nerves develop quite separately from those between the motor nerves and the muscles. The sensory component of the spinal nerve develops from a group of ganglion cells of neural crest origin; central branches of these cells form the dorsal root, which grows into the segment of the neural tube already defined by the outgrowth of the motor root. Together, the dorsal and ventral roots constitute the mixed spinal nerve.
In contrast to the segmental pattern of the nerves, the arteries to the body wall are branches of the aorta that initially pass intersegmentally between the somites (Figure 2-3/5). Despite this, the arteries and nerves later associate in a way that fails to reflect the different patterns of their origins.
The lateral and ventral parts of the body wall are initially unsegmented (see Figure 2-3).
The tissues of these parts develop in the somatopleure, which is formed by the association of the ectoderm and the outer of the two sheets into which the lateral plate mesoderm is split. The inner sheet of the lateral mesoderm is, of course, combined with the endoderm to constitute the splanch- nopleure or gut wall. The separation of these sheets is achieved by the coalescence of initially scattered spaces to form a continuous cavity (Figure 2-5/9). The cavity, known as the celom, is afterward divided to yield the pericardial and pleural spaces of the thorax and the peritoneal space of the abdomen and pelvis. The somatopleure is later invaded by cells that migrate ventrally from local somites. Cells that migrate from the sclerotomes of thoracic somites differentiate to form the ribs and sternum. Cells that migrate from the myotomes of both thoracic and abdominal somites differentiate to form the muscles of the thoracic and abdominal walls. The presence of the ribs ensures that the thoracic wall retains a segmental pattern, which is almost completely lost by the abdominal wall.The embryo is still open ventrally while these events are proceeding. The ventral aspect of the body wall closes only in the final stage of the folding (reversal) process (p. 100) that converts the embryonic disk into a more or less cylindrical body. Ventral midline structures including the sternum and the linea alba—the median connective tissue strip of the abdominal floor—are therefore initially represented bilaterally. The umbilical scar, our “belly button,” betrays the site of final closure of the body wall.
The clinician’s chief interest in the umbilical scar relates to the prevalence of umbilical hernia, a congenital (possibly inherited) defect that frequently occurs in domestic species. Some delay in the closure of the ventral abdominal wall is always necessary to allow for the temporary physiological herniation (p. 145) of a part of the gut into the extraembryonic celom (within the umbilical cord).
Normally the herniated loops of intestine are soon drawn back into the abdomen, and narrowing and, eventually, closure of the peritoneal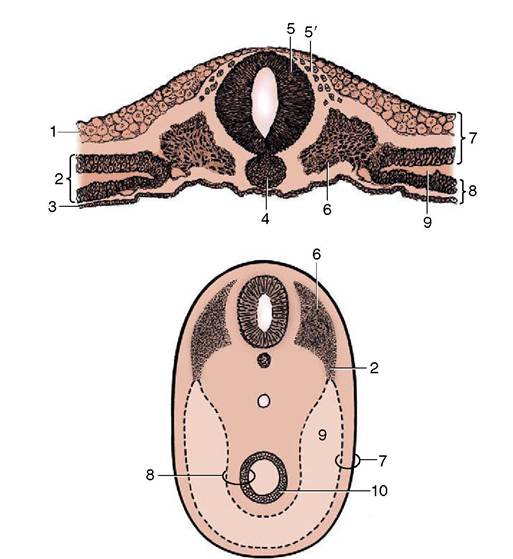
Figure 2-5 Transections of an early discoidal embryo (above) and of an older ventrally closed one to show the splitting of the lateral mesoderm and the development of the celom. 1, Ectoderm; 2, lateral plate of mesoderm; 3, endoderm; 4, notochord; 5, neural tube; 5', neural crest cells; 6, somite; 7, somatopleure; 8, splanchnopleure; 9, celom; 10, primitive gut.
ring at the junction of the intraembryonic and extraem- bryonic parts of the celom then follow. This, in turn, allows the closure of the defect in the mesodermal tissues, creating the umbilical scar. These processes may be faulty. The intestine may fail to complete its return to the abdomen or, once returned, may make a second escape into the umbilical cord through a persistent peritoneal ring and thus be exposed when the cord is ruptured at birth. More commonly, the peritoneal ring closes, but the overlying tissues remain defective and herniation occurs into a protuberant sac formed by stretching of the peritoneum and covering fasciae and skin. Fortunately, umbilical hernia is usually amenable to simple surgical correction.