CARDIOVASCULAR SYSTEM
The heart lies in the midline and slightly to the right and is surrounded by a thin but tough pericardium (Akester 1984), which is attached to the dorsal surface of the sternum, the surrounding airsacs, and liver.
The lungs lie dorsal to the heart and not surrounding it, while the liver lobes cover the base of the heart dorsally and laterally (Smith & Smith 1997).The heart has four chambers with separate right and left sides so that oxygenated and deoxygenated blood never mix. It is quite similar to the mammalian heart except that in birds the ascending aorta curves to the right. In some species, like the chicken and ostrich, the cranial and caudal vena cavae enter first into a sinus venosus before entering the right atrium (King & McLelland 1984; Maina 1996; Rosenthal 1997b). Like reptiles, birds have a renal portal system.
The right atrioventricular valve is structurally unique to birds in that it has no chorda tendinae and forms only a thick, muscular flap of myocardium. The rest of the valves are more similar to mammalian ones, except the left atrioventricular valve is tricuspid (Akester 1984; Smith & Smith 1997). Unlike reptiles, where the muscle tissue of the atrium is continuous with the ventricle, birds have a specialized conducting system of Purkinje fibers which carry the impulse from the atrium to the ventricle. As in mammals, this begins at the sinoatrial node, located in the right atrium (Akester 1984; Smith & Smith 1997; West et al. 1981).
Arterial system
The aortic arch curves to the right and gives rise almost immediately to two very large brachiocephalic trunks which are responsible for distributing about three quarters of the cardiac output to the wings, pectoral muscles, and head (Akester 1971; West et al. 1981). These soon branch into the subclavian arteries, which supply the wings (brachial artery) and flight muscle (pectoral artery), and the carotid arteries which supply the head (Fig.
6.32). The carotid arteries run along a groove at the base of the cervical vertebrae close to the axis of rotation. This prevents movements of the flexible neck arresting blood flow to the brain (King & McLelland 1984; Rosenthal 1997b; Smith & Smith 1997).CLINICAL NOTE
Unlike mammals, the heart is obscured at its base by liver lobes rather than the lungs. This gives an hourglass appearance to the cardiac and hepatic shadow on ventrodorsal radiographs in many psittacine species (Figs. 6.30 and 6.31). However, macaws have a relatively small liver, which might lead to an over diagnosis of cardiomegaly (Smith & Smith 1997).
114
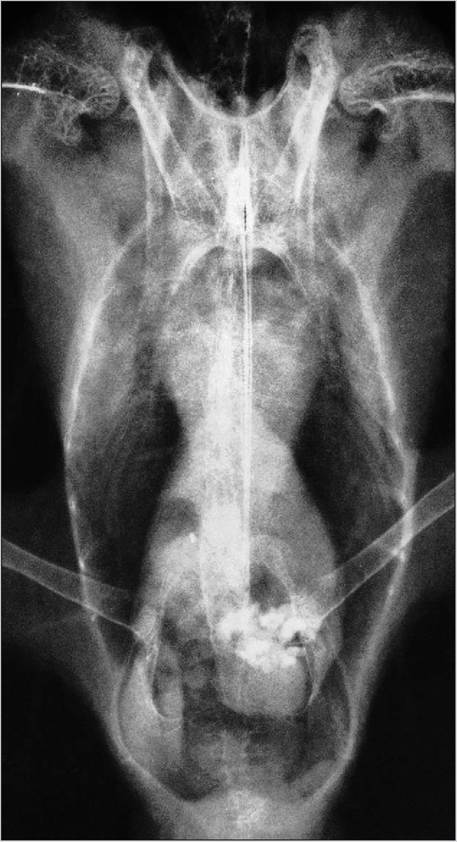
Figure 6.30 • Ventrodorsal radiograph showing normal cardiac and hepatic silhouette in a cockatiel (Nymphicus hollandicus). Grit is visible in the gizzard which lies at the level of the acetabuli.
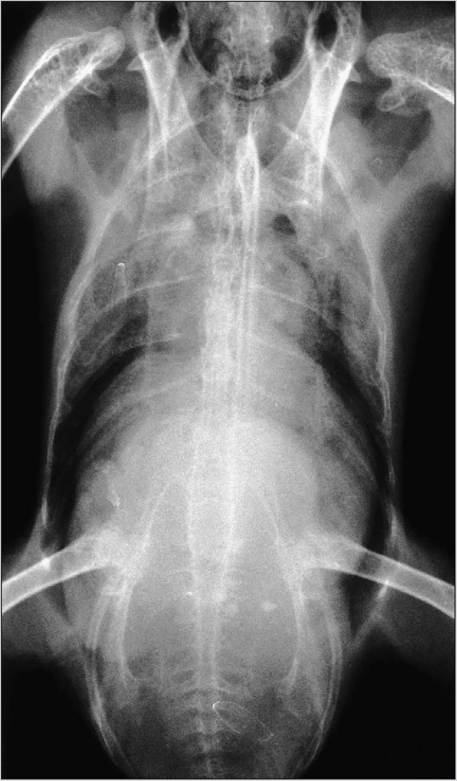
Figure 6.31 • Radiograph showing hepatomegaly secondary to lymphoma in an African gray parrot (Psittacus erithacus).
CLINICAL NOTE
The large brachiocephalic trunks can often be seen end on in ventrodorsal radiographs of large birds and must not be confused with fungal granulomas (Smith & Smith 1997).
The pelvic limb is supplied by the external iliac artery and the ischiatic artery that meets the femoral artery at the stifle to form the popliteal artery. It is this artery that forms an arteriovenous network of vessels (rete mirabile) in many wading birds. This aids in heat exchange by warming cold blood from the extremities (Smith & Smith 1997).
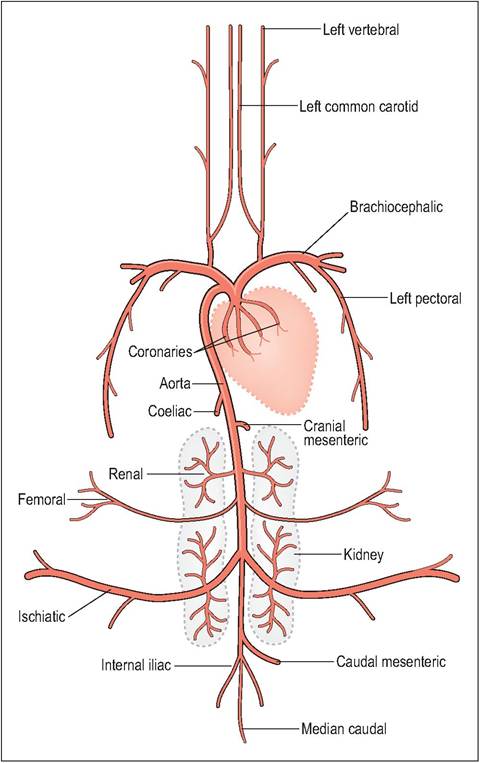
Figure 6.32 • The major systemic arteries in the bird.
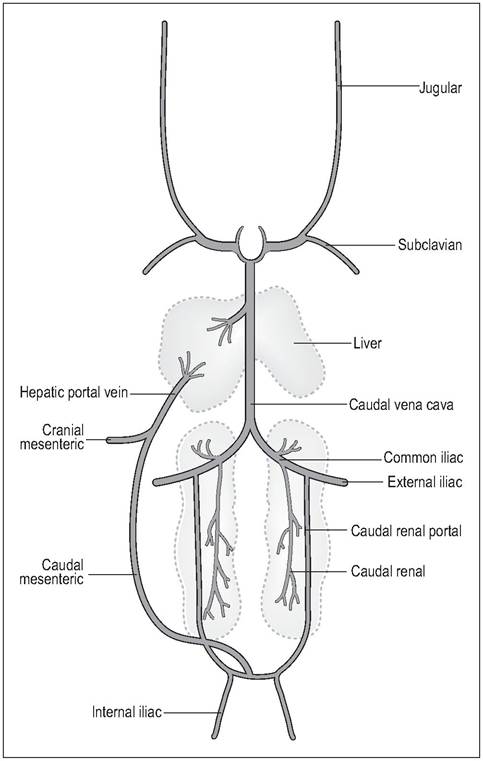
Figure 6.33 • The major systemic veins of birds.
Venous system
The cranial vena cavae are paired and receive blood from the neck and head via the jugulars, and from the wing and breast via the subclavian veins. At the angle of the jaw there is a transverse anastomosis between the two jugulars which allows blood to bypass from one side to another in case one vein gets compressed during neck movement. This anastomosis slopes caudally towards the right jugular and this may explain why, in most species, the right jugular is much larger than the left (Akester 1971; West et al. 1981). In some small cage birds the left jugular is lost altogether (Evans 1996; King & McLelland 1984).
Venous blood from the pelvic limbs, lower body, and hindgut enters the renal portal system, which then joins the caudal vena cava. The main bulk of the gastrointestinal tract, pancreas, and spleen drains into the hepatic portal vein and the liver. Unique to birds is the large caudal mesenteric or coccygeal mesenteric vein, which drains the hindgut mesentery and connects the hepatic portal vein to the renal portal vein. As blood can flow along this vein in both directions flow can be switched between the kidneys and the liver (Akester 1971) (Fig. 6.33).
Renal portal system
In birds, venous return from the pelvic limbs goes through the kidneys before reaching the caudal vena cava, contributing about two thirds of renal flow (Smith & Smith 1997). This is controlled by portal valves at the junction of the common iliac and renal veins (Akester 1971; West et al. 1981). These are unique in that they contain innervated smooth muscles. Normally, these valves are closed to direct blood through the kidney where they take part in tubular secretion (see Urinary system). In times of stress adrenaline opens these valves to allow the blood to bypass the kidneys completely. It can then take a short cut via the coccygeal mesenteric vein to the liver or directly into the caudal vena cava to the heart (Akester 1971, 1982; Smith & Smith 1997; West et al.
1981).Venepuncture sites
(See Echols 1999.)
Three large peripheral veins are suitable for avian blood sampling:
116
The deep ulnar vein (basilic) - This becomes cutaneous at the elbow on the ventral aspect of the wing. Hematoma formation is always a problem so digital pressure must be applied immediately post sampling. Right jugular vein - This is the least prone to extravasation post sampling. The vein lies in a natural featherless tract (aptyeria) so can be easily visualized in most birds. However, Columbiformes and Anseriformes lack apteria, making visualization more difficult (Echolls 1999) (Fig. 6.34).
Medial metatarsal - This is less likely to form a hematoma, owing to the scales of the leg. It is a useful venipuncture site in waterfowl, although feathering on the leg and the thick skin can make this vessel harder to identify.
Toe-nail clipping - Small samples can also be taken by this method but it can be painful and the nail must be cleaned well to avoid contamination from urates.
Cardiac physiology
In order to be able to fly birds have high oxygen demands, so consequently the avian heart is 50 to 100% larger than in mammals of the same size (Maina 1996; Rosenthal 1997b; Smith & Smith 1997). The size of the heart also depends on the amount of aerobic energy each species expends. For example, a large bird like a swan will have a proportionately smaller heart for its size than a racing pigeon (Maina 1996; Smith & Smith 1997). Birds also have a far greater exercise capacity than humans: at maximum exercise, the cardiac output in a flying budgie is seven times greater than that of a man or dog (King & King 1979; King & McLelland 1984; Maina 1996).
In order to pump large volumes of blood to the wings, head, and flight muscles, birds have a much higher cardiac output than mammals. This is achieved by a high stroke volume, fast heart rates (150-350 resting), and slightly lower peripheral resistance. They also have stiffer arteries to improve blood flow and maintain a high blood pressure, ranging
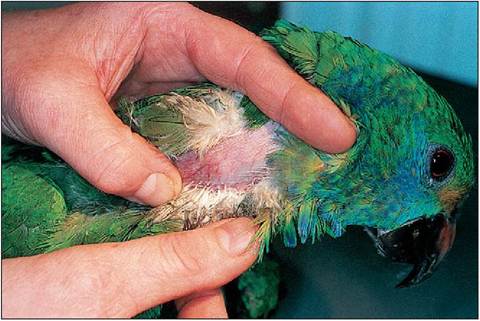
Figure 6.34 • Blood sampling from the right jugular vein of an Amazon parrot.
GENERAL INTEREST
The hummingbird has the largest heart of all, relative to its size, in order to sustain the aerobic demand of hovering.
In contrast to the mouse (where the heart accounts for 0.5% of its body mass) its heart represents 2.4%. When hovering it can reach a heart rate of over 1000 beats per minute (King & McLelland 1984).
from 108-250 mmHg (compared to an average of 150 mmHg in humans). Turkeys have the highest pressure of all vertebrates, being as high as 350 mmHg (Akester 1984; Maina 1996). The consequences of this high pressure can, however, mean that aortic rupture, heart failure and hemorrhage are a common cause of death in stressed avian patients (Rosenthal 1997b; Welty 1982b).
CLINICAL NOTE
Grossly, the major avian arteries appear white due to the collagen fibers of the tunica adventitia. These fibers produce the stiff arteries that help combat the high blood pressure and improve blood flow. The downside of this is that birds are susceptible to atherosclerosis of the aorta and brachiocephalic trunk. This is commonly seen in aged Amazon parrots (Rosenthal 1997b).
Diving birds
Unlike reptiles, birds cannot shunt blood away from the lungs while diving because the ventricles are completely divided. Instead, diving birds use selective vasoconstriction, bradycardia, and a drop in cardiac output. Blood flow is diverted from the skin, muscles and gastrointestinal tract to the brain, sensory organs, and myocardium to protect them from hypoxia during a dive (Akester 1984; Fowler 1986; Rosenthal 1997b).
Blood components
Blood volume varies from 5 to 13% of body mass. The blood cells are made up of nucleated erythrocytes, thrombocytes, lymphocytes, monocytes, heterophils, eosinophils, and basophils.
Erythrocytes
The avian erythrocytes are oval in shape and, being nucleated, are usually larger than those in mammals (Fig. 6.35). Flightless birds like the ostrich have the largest erythrocytes whereas the more higher evolved good fliers, like passerines, tend to have smaller erythrocytes (Maina 1996; Welty 1982b).
Small erythrocytes produce a greater combined surface area and more efficient gas exchange, thus allowing for higher oxygen absorption and hence a metabolic rate that is 10 times that of mammals (Hodges 1979; King & McLelland 1984).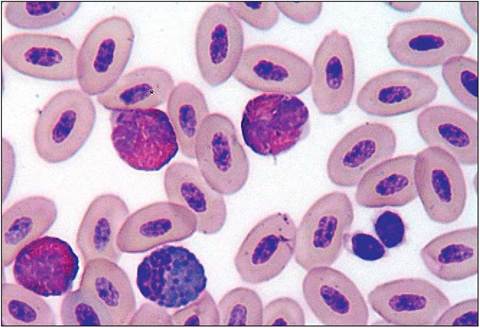
Figure 6.35 • Blood smear from an African gray showing nucleated avian erythrocytes; three heterophils, one eosinophil and two thrombocytes.
The avian erythrocyte does not necessarily contain more hemoglobin than mammals but species that fly at high altitudes have a higher hemoglobin affinity (Butler & Bishop 2000; Welty 1982b).
CLINICAL NOTE
EDTA causes lysis of red blood cells in certain species, like the ostrich, crow, and crane. Avian leukocytes are very fragile so particular care must be taken when blood smears are made. The use of cover slip smears may avoid cell disruption. Smears should be made immediately after collection because blood coagulation takes place faster in birds (30-60 s) than in mammals.
GENERAL INTEREST
The ability to absorb more oxygen means that birds can fly at high altitudes. The Bar-headed goose (Anserindicus) migrates over Mount Everest and the Himalayas and has the amazing ability to fly from sea level to an altitude of 9200 m without having to acclimatize (King & King 1979; Scheid & Piiper 1972).
Control of erythropoiesis
Erythropoiesis takes place in the bone marrow. The avian erythrocyte has a short life span compared to that in mammals, presumably due to their high metabolic rate and body temperature. For example, duck and pigeon erythrocytes last 42 and 35-45 days, respectively, compared to 90-120 and 45-50 days in humans and rats (Freeman 1971; Maina 1996). This short life span means that persistent anemias should be investigated thoroughly.
Blood loss
Birds (especially flying ones) are better able to tolerate blood loss than mammals, with the pigeon being able to lose 8% of its body mass from hemorrhage. This rapid restoration of blood volume may be due to pronounced vasoconstriction in skeletal muscles raising the blood pressure and to large capillary beds that ensure rapid absorption of tissue fluid (Smith & Smith 1997).
Thrombocytes
Thrombocytes are fragile, nucleated cells analogous to the mammalian platelet, although they originate from a stem cell and not from megakaryocytes like platelets (Freeman 1971; Powers 2000). Although avian thrombocytes are involved in the clotting process they have little thromboplastin and so do not appear to trigger the intrinsic clotting process (Archer 1971; Hodges 1979; Powers 2000). Instead, unlike mammals, it is the extrinsic thromboplastin shed by the damaged tissue that plays a more major role in clotting. This reliance on extrinsic clotting pathways may exist to prevent exsanguination in animals with such high blood pressure and rapid heart rates (Powers 2000).
Lymphocytes
T lymphocytes are produced by the thymus. They have a long life span and develop cellular immunity. B lymphocytes produced by the Bursa of Fabricius are involved with humoral cell responses and have a short life span.
Monocytes
These are rarely seen in peripheral blood smears, so low or zero counts are common. They can be elevated with chronic infections like psittacosis, tuberculosis, and mycotic infections.
Heterophils and eosinophils
Heterophils are similar to mammalian neutrophils, having a polymorphic nucleus. They are the equivalent of neutrophils but stain acidophilic (Hodges 1979). Eosinophils account for about 2% of total leukocytes (Freeman 1971); their function is uncertain but may be associated with tissue damage or parasitic disease.
KEY POINTS
• Avian heart is larger for body size than mammals
• Aorta arches to the right in birds. Brachiocephalic vessels are very large as they supply pectoral arteries for flight
• Heart and liver form an hourglass appearance on ventrodorsal radiographs in many psittacines
• Renal portal vein
• Nucleated erythrocytes
• Higher blood pressure means greater tendency to hemorrhage, although birds are surprisingly tolerant of blood loss
• EDTA lyses red blood cells in some species so a fresh blood smear is always advisable