SKELETAL SYSTEM
There are two main subclasses of bird in existence today and these are based on the anatomical structure of the sternum. These are the ratites, which include the flightless emu, ostrich (Fig.
6.3), and kiwi, and the carinates, which include the rest of avian species (8616 species). The largest living carinate is the Andean condor (Vultur gryphus) which has a wingspan of 3 m and weighs 15 kg.Ossification of bones
Cortical bone is similar in both sexes but in the female the medullary cavity is very labile and is the most important calcium reserve for the egg (Taylor et al. 1971; Tully 2002). Prior to laying, medullary bone draws calcium from the alimentary tract to calcify the medullary cavity (Johnson, AL 2000). Bony trabeculae are laid down from the endosteum and the total skeleton increases by about 20%. This phenomenon, called polyostotic hyperostosis, is visible radiographically and is followed by bone resorption once the eggshell is calcified (Fig. 6.62).
Like mammals, birds ossify their skeleton on a cartilaginous model although secondary centers of ossification are lacking. The cortex is relatively thin but the medulla is bridged by numerous trabecular struts to add extra strength (Evans 1996; Taylor et al. 1971; Tully 2002).
CLINICAL NOTE
The thin avian cortices and internal bone struts mean that bones can splinter very easily, making orthopedic surgery a challenge at times (Orosz 2002).
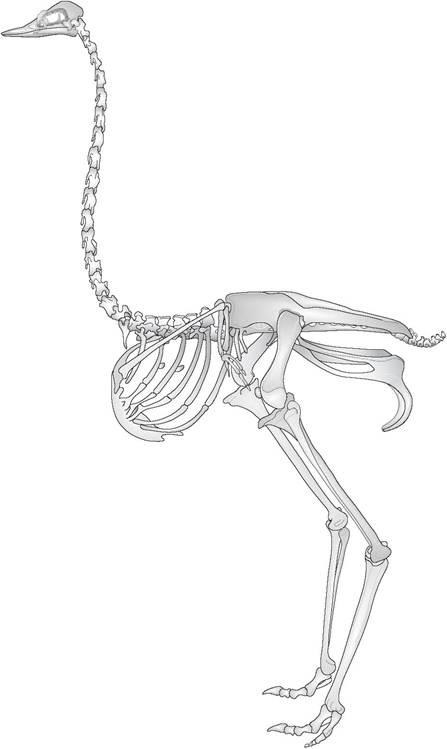
Figure 6.3 • The skeleton of a ratite (ostrich, Struthio camelus), showing flat sternum, rudimentary pectoral girdle and vestigial wings. Unlike other avian species the ostrich also has a pubic symphysis which may be an adaptation to support the heavy mass of the viscera.
Skeletal modifications for flight
Birds have a lightweight fused skeleton.
For example, the skeleton of a pigeon is 4.4% of body mass compared to that of a rat's skeleton which is 5.6% (King & King 1979; Maina 1996) (Fig. 6.4).■ The avian forelimb is modified into a wing while the bill and neck are modified for food prehension. The manus is tapered and fused to hold the primary feathers (Figs. 6.5 and 6.6).
■ Many bones of the backbone and limbs are fused to form a rigid and strong but light framework. The fused rib cage helps resist the twisting and bending of wings in flight while the rigid pectoral girdle acts like a wing strut. A fused tail vertebra (pygostyle) provides a short tail for steering and maneuverability. The sternum is keeled (carinate) to hold the muscles of flight (King & Custance 1982; King & King 1979).
■ The airsacs extend into the medullary cavity of the major bones, such as the humerus, coracoid, pelvis, sternum, and vertebrae. They are most developed in the good fliers to help in weight reduction. In some birds the femur, scapula and furcula are also pneumatized but this does not tend to happen to the distal bones. The skull also consists of a honeycomb of air spaces with delicate spicules for support (Koch 1973; Maina 1996).
■ The supracoracoid muscle lifts the wing by passing from its ventral attachment on the sternum through the triosseal foramen to insert on the dorsal humerus. This keeps all the heavy flight muscles along with the muscular gizzard situated ventrally at the bird's center of gravity (King & Custance 1982).
Skull
The cranial bones of the skull are fused to form a rigid, but lightweight, box with large orbits separated by a thin, bony
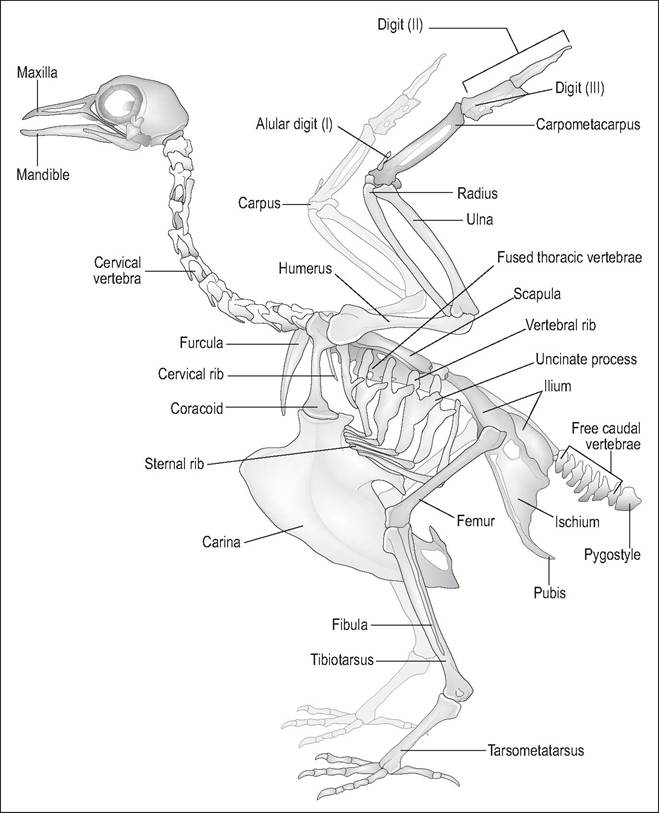
Figure 6.4 • The skeleton of a carinate, the pigeon, showing modifications for flight: large keeled sternum for flight muscles, fused thoracic vertebrae, strong pectoral girdle for bracing the wings and large feet to withstand the concussion of landing.
interorbital septum (Figs. 6.10-6.12). The brain has been pushed caudally and ventrally into the occipital region and lies at a 45-degree angle tilt. A single occipital condyle articulates with the atlas, allowing birds to rotate their neck to an angle of 180 degrees (Dyce et al. 2002; Koch 1973).
Within the rostral skull there are large areas of honeycomb pockets of air or sinuses that are especially prominent in flighted birds. The infraorbital sinus has many diverticula and is very well developed in psittacines. Diving birds and birds that peck at hard objects (e.g., woodpeckers) lack these pneumatic zones in order to help the skull withstand more concussive force.
Cranial kinesis
Birds, especially psittacines, have a highly mobile kinetic skull. This means that, unlike mammals that can only move their bottom jaw, they are also able to move their maxillary jaw (upper beak). This wide gape is achieved by an elastic hinge at the rostral skull that allows the bones to bend without disturbing the cranium (Fig. 6.7). In psittacines, this elastic hinge is replaced by an articular craniofacial joint, allowing parrots even more flexibility of movement (Evans 1996; Quesenberry et al. 1997).
The mobile quadrate bone also plays a major role in skull kinesis. This bone not only articulates with the cranium but also with the premaxilla via two rodlike thin bones called the jugal arch (precursor of the zygomatic bone) and pterygoid-palatine bone. When the jawbone is lowered the quadrate bone pushes these two bones rostrally to elevate the upper jaw, allowing the bird a wide gape (Evans 1996; Maina 1996).
Premaxilla
The upper jaw is derived from the premaxillary and nasal bone and a small part of the maxillary bone. This is very thin and lightweight, owing to the diverticula extending from the infraorbital sinus. The kinetic movement of this upper jaw is either prokinetic or rhynchokinetic. In prokinetic birds, like psittacines and chickens, the upper jaw moves as a unit and the nasal openings are small and oval (Fig.
6.8). In rhyn- chokinetic birds (e.g., pigeons and waterfowl) only the rostral part of the upper jaw moves and the nasal openings are elon-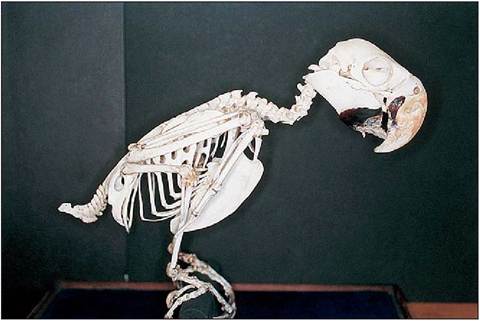
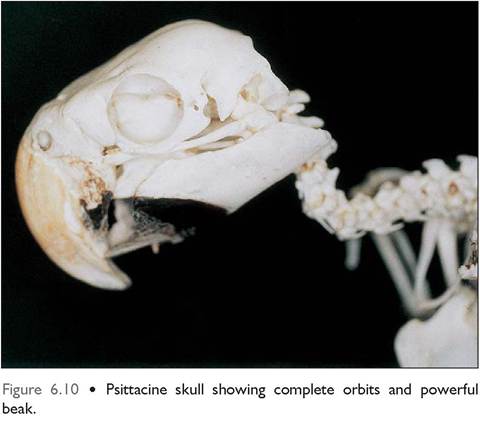
Figure 6.5 • Skeleton of Blue and gold macaw (Ara ararauna). Note the s-shaped neck, the powerful beak (the lower beak is damaged in this specimen) and complete orbit of the psittacine.

Figure 6.6 • Blue and gold macaw (Ara ararauna).
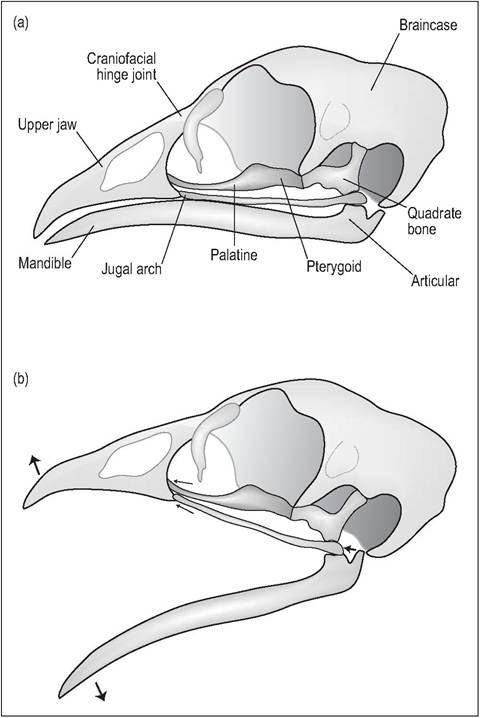
Figure 6.7 • Cranial kinesis.
(a) Skull of domestic fowl (Gallus gallus) with mouth closed.
(b) Open mouth view. When the jawbone is lowered the quadrate bone pushes the jugal arch and pterygoid-palatine bone rostrally to elevate the upper jaw, allowing the bird a wide gape.
gated and slitlike (King & McLelland 1984; Quesenberry et al. 1997) (Fig. 6.9).
Mandible
The mandible in birds consists of five small bones which fuse caudally with the articular bone. The most rostral bone is the dentary bone and this forms a fully ossified mandibular symphysis. The others are the surangular, angular, splenial and prearticular. Caudally, the articular bone articulates with the quadrate bone. In mammals these two bones have evolved into the auditory bones incus and malleus.
KEY POINTS
• Avian skull is highly kinetic
• Movable quadrate bone allows wide gape
• Single occipital condyle so can rotate head 180 degrees
• Well-developed sinuses
• Psittacines have synovial joint at craniofacial maxillary hinge for greater gape
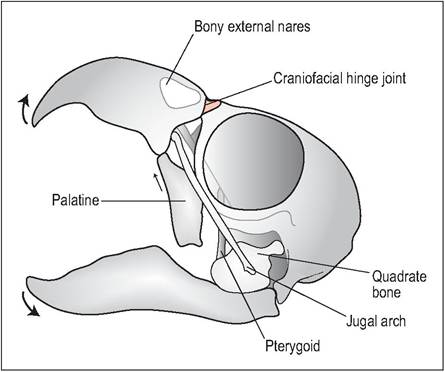
Figure 6.8 • Prokinesis.
The upper jaw moves as a unit and the nasal openings are small and oval. This is particularly developed in psittacines, which have a craniofacial hinge.
id="Picutre 124" class="lazyload" data-src="/files/uch_group31/uch_pgroup304/uch_uch7234/image/image124.jpg">
Figure 6.9 • Rhynchokinesis.
In birds like pigeons and waterfowl only the rostral part of the upper jaw moves and the nasal openings are elongated and slitlike.
Axial skeleton
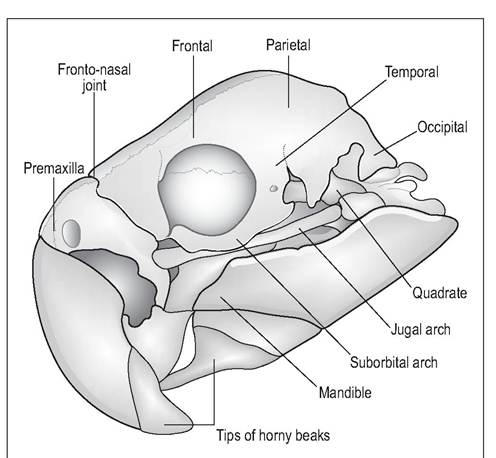
Figure 6.11 • Psittacine skull.

Figure 6.12 • Skull of eagle owl (Bubo bubo) demonstrating large incomplete orbits and powerful hooked beak for tearing at prey.
fine control of the tail feathers. The number of vertebrae
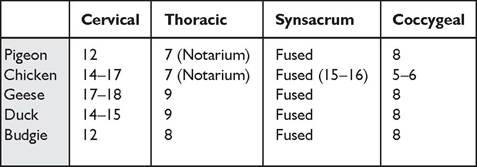
Birds have epaxial muscles dorsally and hypaxial muscles ventrally along the vertebral column. These are most developed in the neck for preening and the apprehension of food. The tail muscles are also well developed for can vary widely according to species, as can be seen in Table 6.2.
Cervical vertebrae
The forelimbs are modified for flight so the neck and beak play a larger role in grooming and manipulation of objects. In general, necks tend to be longer in waterfowl, which need to be able to reach the uropygial gland for preening.
The cervical vertebrae are long and flexible (numbering from 8 in small birds to 25 in swans) with highly mobile, saddle shaped, articular surfaces enabling it to adopt the
Table 6.2 Number of vertebrae in common avian species (Evans 1996; Koch 1973)
sigmoid shape bend. Rostrally, the atlas articulates in a ball and socket joint with a single occipital condyle giving head movement great flexibility (Maina 1996). This allows the bird to rotate its head to compensate for poorly developed eye muscles. The caudal cervical vertebrae have rudimentary ribs which are the site of attachment of the cervical muscles (King & McLelland 1984; Koch 1973).
acting like a protective brace around the chest and heart. These are particularly well developed in diving birds such as guillemots to help the thorax withstand the pressure of a dive.CLINICAL NOTE
CLINICAL NOTE
The soft tissues of the neck, the esophagus, and trachea are shorter than the cervical vertebrae, so it is impossible to stretch a bird's neck out completely. This sigmoid neck acts like a spring to protect the head and brain from concussive forces while landing. Hence the elegant s-shaped neck of the swan (Koch 1973) (Fig. 6.13).

Figure 6.13 • Mute swan (Cygnus olor) showing ‘s'-shaped neck.
In psittacines the last rib does not possess an uncinate process so this provides a useful surgical and laparoscopic landmark (Quesenberry et al. 1997).
Synsacrum
This contains from 10 to 23 vertebrae and is the fusion of caudal thoracic, lumbar, sacral, and caudal vertebrae. It supports the pelvic girdle and hence the bird’s entire mass (Fig. 6.15).
Caudal vertebrae
The tail is short and the last vertebrae are fused into a single, flattened bone called the pygostyle, which supports the tail feathers (Fig. 6.14). This is most highly developed in birds that use their tail for climbing and support. Well-developed muscles are present to help control the pitch during flight (King & King 1979; King & McLelland 1984).
Sternum
This is much more extensive than in mammals, being a ventral plate of bone providing protection. The keel bone (carina) provides the main attachment for the flight muscles. It is most developed in sophisticated fliers like swifts and hummingbirds and least developed in the flightless ratites, which have a flat and raft-like sternum (Bezuidenhout 1999; King & McLelland 1984; Maina 1996).
Thoracic vertebrae
In birds, much of the lower vertebrae can be fused to confer rigidity on the skeleton for flight. Many species, like chickens, hawks, and pigeons, have the first 3 to 5 thoracic vertebrae fused into a single bone, the notarium, which provides a rigid beam to support flight (King & McLelland 1984; Koch 1973). This is followed by the only mobile vertebra of the trunk. This can be a weak link because, when ventrally displaced, it causes spondylolisthesis or “kinky back” in broilers (Dyce et al. 2002). Budgies have mobile thoracic vertebrae at T6-7 (Evans 1996).
Thoracic vertebrae vary in number from 3 to 10 and can be identified by ribs, which articulate with the sternum. Some cranial and caudal ribs lack a sternal attachment but have a ligamentous attachment instead. External and internal intercostal muscles lie between each rib. A unique feature of avian ribs is a backward-pointing process, the uncinate process, which extends caudodorsally from every rib. This provides attachment for muscles which extend ventrocaudally to the rib behind, adding strength to the thoracic wall and
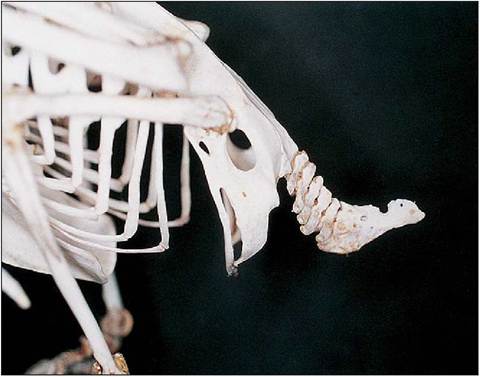
Figure 6.14 • Lateral view of psittacine pelvis, caudal vertebrae and pygostyle.
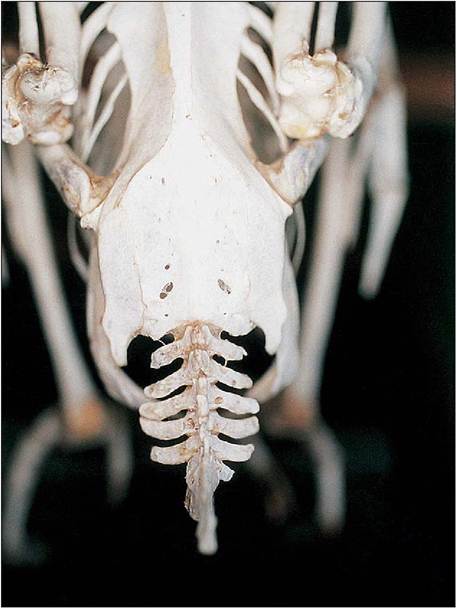
Figure 6.15 • Dorsal view of psittacine synsacrum, pelvis, caudal vertebrae and pygostyle.
The caudal portion of the sternum is often notched or perforated by windows. These are areas of the sternum that have failed to ossify and are lined by fibrous membranes. They are most developed in poor fliers like poultry (Dyce et al. 2002; Koch 1973).
Appendicular skeleton
Pectoral girdle
This consists of the clavicle, coracoid, and scapula, all of which articulate proximally to form the triosseal foramen. Distally, the scapula and coracoid also form the glenoid cavity, which is shallow and directed laterally to allow the wings adduct and abduct for flight (Evans 1996) (Fig. 6.16).
Clavicle
Along with the coracoid, this acts to brace the wing and is the proximal attachment of the pectoralis muscle. These are rudimentary in many Psittaciformes and Strigiformes (Evans 1996; King & McLelland 1984). In many species the two clavicles are fused into the furcula (wishbone) for extra strength. This unites both shoulder blades and has a springlike function (Koch 1973).
Coracoid
This is a short, strong bone which extends from the sternum and acts like a wing strut in preventing the wing muscles from compressing the thorax on the downstroke (King & McLelland 1984). It lies deep below the cranial edge of the pectoralis muscle and lateral to the jugular veins and subclavian and common carotid arteries.
Scapula
This is a long blade-like bone whose length varies with the flying strength of each species. It lies parallel to the backbone and extends as far caudally as the pelvis.
Pelvic girdle
The pelvic girdle rotates backward and functions for perching and locomotion like running and swimming. It is made up of a fusion of the ischium, pubis, and ilium and is rigidly
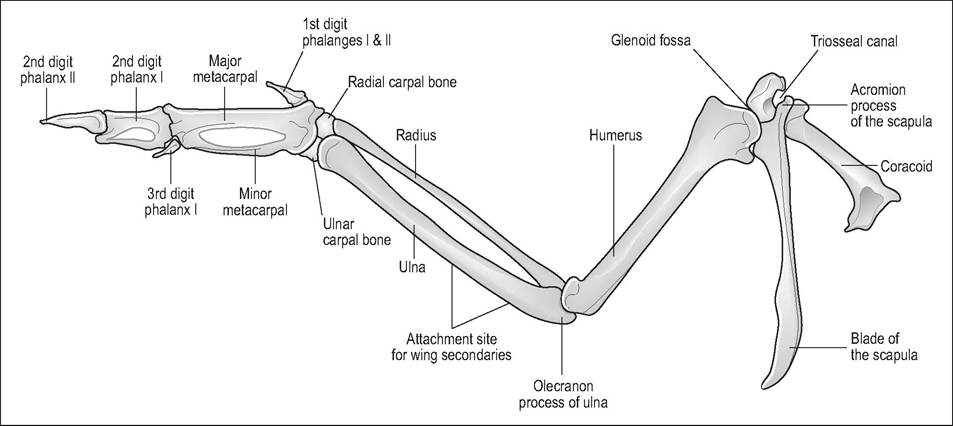
Figure 6.16 • Ventral view of pectoral girdle and left wing.
CLINICAL NOTE
As the coracoid bone acts as the brace between wing and sternum it is commonly damaged during wing trauma. If the wing is drooping and the humerus feels intact, radiography may be necessary to check the coracoid bone for fractures.
fused to the synsacrum at the ilium. In most species it is incomplete ventrally, presumably to allow for passage of eggs through the pelvic canal (King & Custance 1982). The exceptions are the ostrich and rhea, which have a pubic symphysis that may be an adaptation to support the heavy mass of viscera (Bezuidenhout 1999; Evans 1996; Koch 1973; Maina 1996; Raikow 1985) (Fig. 6.17).
The acetabulum is deep and, as there is a strong femoral attachment, luxation is uncommon (Quesenberry et al. 1997). The trochanter of the femur also articulates with the antitrochanter of the ilium and this additional joint enables birds to perch on one leg very easily (Maina 1996). Ventrally there are bilateral renal fossae for the caudal division of the kidneys.
Wings
The wing skeleton consists of humerus, radius, ulna, carpal bones, carpometacarpus, and three digits (Orosz et al. 1992).
Humerus
The humerus is a short bone that lies against the body wall when the wing is folded. Proximally, it has a well-developed pectoral crest where the pectoralis muscles insert. The biceps brachii lies along the cranial humerus and serves to flex the wing while the triceps brachii lies caudally and serves to extend the wing. The clavicular airsac extends into the medullary cavity, making this bone pneumatic.
Radius and ulna
These are long bones that lie parallel to each other. In birds, these bones do not twist and the ulna, which lies caudally, is the bigger bone. Small bony knobs can be seen on the caudal aspect where the secondary flight feathers insert.
Manus
The wrist and hand are severely reduced to provide a strong base for the attachment of the primary flight feathers. Only
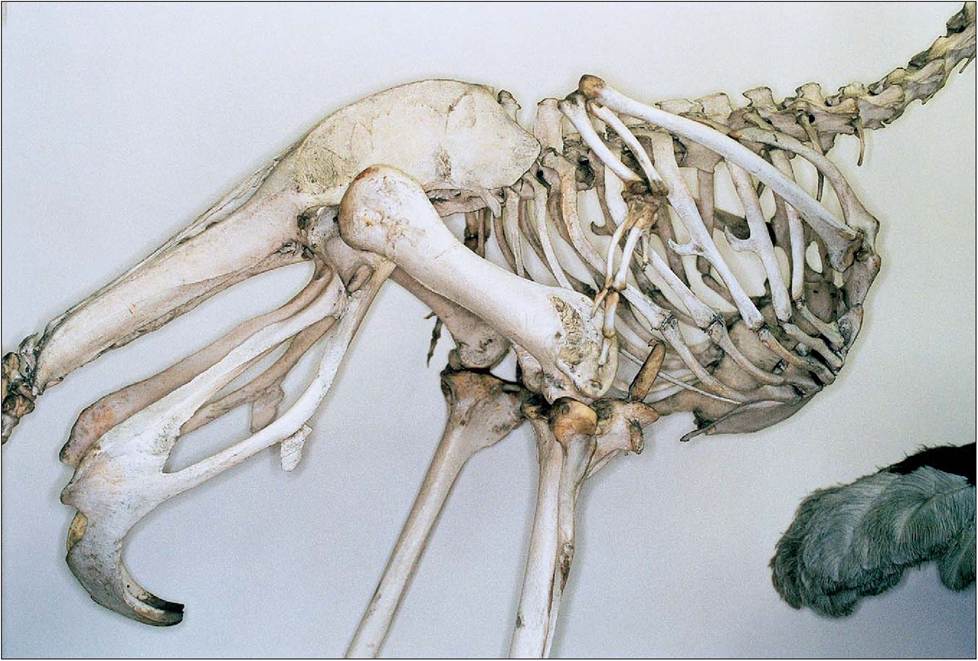
Figure 6.17 • Sternum and pelvis of ostrich (Struthio camelus). The pubic symphysis may be an adaptation to support the heavy mass of viscera in these species.
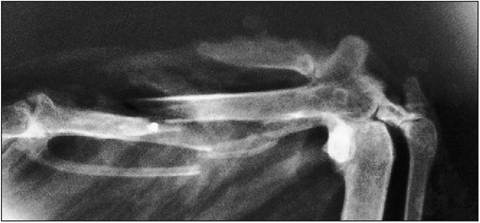
Figure 6.18 • Radiograph of distal wing of Buzzard (Buteo buteo) with major and minor metacarpals fractured by lead shot.
CLINICAL NOTE
The secondary flight feathers insert onto the caudal ulna bones. The distal ulna is also the site for placement of intraosseous catheters.
the ulnar and radial carpal bones remain to form the proximal carpus, which is then attached to the Carpometacarpus. There are three digits. The first digit is the alular digit and can have 1 or 2 phalanges. This forms the “bastard wing” and is more mobile than the rest. Feathers attached to this digit are raised during flight to prevent stalling at slow speeds (Orosz 2002).
The major metacarpal bone articulates with the major or second digit, which has two phalanges, while the minor metacarpal bone articulates with the minor or third digit (Fig. 6.18). The carpus and elbow joints are restricted only to extensions and flexion in order to provide a rigid wing surface. They flex simultaneously to fold the wing (Orosz 2002; Orosz et al. 1992).
Pinioning
This is when the manus is cut off at the carpus on one wing to prevent flight. It is frequently done in waterfowl collections at between 4 to 10 days of age. The alular digit is preserved and a cut is made through the two metacarpals as close as possible to the carpus (Dyce et al. 2002; King & McLelland 1984).
Wing types
Four basic wing types can be identified: elliptical, broad, narrow and tapered (Fig. 6.20, Table 6.3). Passerines and pigeons maneuver through undergrowth and have short, broad elliptical wings with rapid beats. They have a large alula and wing slots to prevent stalling at low speeds. Eagles and vultures have a long broad wing that enables them to soar at low speeds. These birds have a large alula and wing slots. Ocean-going birds like gulls and gannets have a long, narrow, and pointed wing with no wing slots, allowing them to soar at high speeds. Swifts and falcons have a tapered wing with no wing slots, which allows them to fly with rapid wing beats at high speeds (King & King 1979; King & McLelland 1984; Maina 1996).
HOW DO BIRDS FLY?
Contrary to popular belief, birds fly in a very different manner to airplanes as their wings provide not just the lift but the propulsive force as well. It is the primary wing feathers which provide most of this propulsion and the secondaries that provide the lift, which is why pinioning of waterfowl is so successful. Smaller birds become rapidly airborne, but large birds like the swan have to “taxi” along the water before they can take off (Spearman & Hardy 1985).
The dorsal surface of the bird's wing is convex and the ventral surface is concave. The leading edge is thickest where bones and muscles provide strength and tapers caudally to the trailing edge. Air traveling over the dorsal wing has to travel further than air passing ventrally and this increased speed produces low pressure dorsally. As the air passes over it generates an upward force called lift. This aerodynamic force is what enables the bird to rise into the air and defy gravity (Spearman & Hardy 1985).
If the leading edge of the wing is tilted the lift force is increased. Wing control is achieved by changes in position of the wings and slots between the feathers (Fig. 6.19). To avoid stalling at low speeds, slots are formed by the separation of the primary feathers and the alular or bastard wing. These slots let through some air to maintain a smooth stream on the upper surface so as to increase lift.
A drag force opposes the forward motion and reduces the lift. The aerofoil design of the bird's wing provides maximum lift and minimum drag.
107

Musculature
The main bulk of muscles in flying birds is concentrated in the pectoral muscles, which can represent up to one third of body mass. This concentration of the muscle mass ventrally, near the center of gravity, provides stability for flight.
Unlike mammals, birds have a very sparse dorsal musculature as the axial skeleton is so tightly fused together that little muscle support is necessary. The muscles of the limbs and wings are also sparse, with many lightweight tendons
| Table 6.3 Wing shape in relation to type of flight (King & McLelland 1984) | ||
| Wing type | Type of species | Type of flight |
| Elliptical, short & broad | Passerines, pheasants, | Rapid wing beat |
| (wing slots) | pigeons | |
| Long and broad (wing | Vultures, eagles, | Low speed |
| slots) | pelicans | soaring |
| Tapered (no wing slots) | Swifts, falcons, | Rapid, high-speed |
| hummingbirds, terns | wing beat | |
| Long, narrow and | Albatrosses, gulls, | High-speed |
| pointed (no wing slots) | gannets | soaring |
Birds
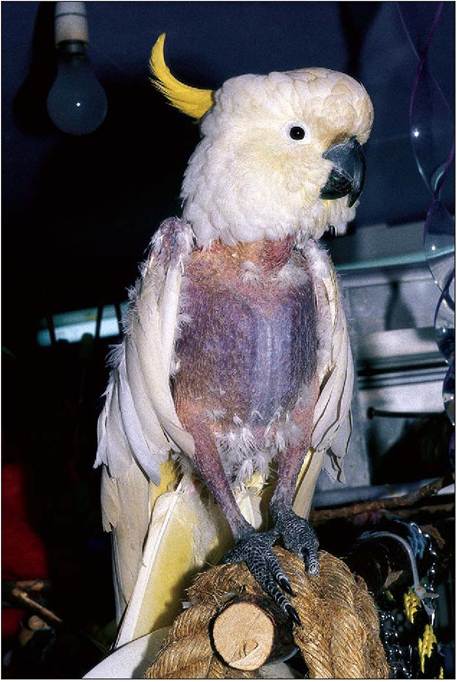
Figure 6.21 • Flighted birds like this Sulphur-crested cockatoo (Cacatua sulphurea) have a high level of myoglobin in the pectoral muscles. The breast muscles appear red in contrast to the white breast muscles of the flightless chicken. (Photo by Claire Nuttall.)
108
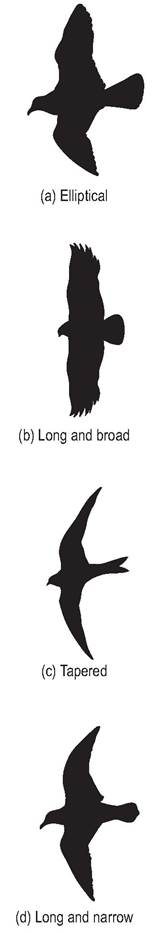
Figure 6.20 • Comparison of four basic wing types.
(a) Elliptical
(b) Long and broad
(c) Tapered
(d) Long and narrow
(Koch 1973). Birds like swifts, which rarely land, have practically redundant leg musculature, while the flightless ostrich has large leg muscles to help them run at high speeds.
Avian muscles have a mixture of both white and red muscles. Red muscle appears red due to the abundance of the oxygen-carrying pigment myoglobin and has an extensive blood supply. As these utilize aerobic metabolism, which can work for long periods, it is found in flight muscles. White muscle is powered by anaerobic metabolism, which is good for rapid contractions, but it tires easily due to lactic acid build up (King & McLelland 1984).
The function of the muscle can be identified by its color (Fig. 6.21). For example, we all know the flightless chicken has white breast muscle and a darker colored drumstick. This is because the gastrocnemius leg muscles contain ten times more myoglobin than the breast. The flighted pigeon, conversely, has pectoralis muscle containing 40 times more myoglobin than chicken muscle (Dyce et al. 2002).
Muscles of flight
The muscles of flight are the pectoralis and the supracoracoid muscles and can represent about 20% of body mass. The pectoralis provides the powerful down stroke and the supracoracoid provides the upstroke (Raikow 1985). Flightless birds have a poorly developed keel bone and, hence, pectoral muscles.
The pectoral muscles extend from the sternum to insert on the medial pectoral crest of the humerus. In most birds like Psittaciformes, Falconiformes, and Columbiformes, the superficial pectoral muscles form the main pectoral mass as they create the downstroke and are consequently essential for flapping flight. Birds like vultures and ocean going seabirds, which glide and soar over long distances, have a well- developed deep pectoral muscle to help brace the wing against the force of wind and keep it motionless while gliding (King & King 1979; Orosz 2002; Orosz et al. 1992).
The supracoracoid muscle also attaches to the ventral sternum beneath the pectoral muscles (Fig. 6.22). Its tendon then runs dorsally through the triosseal canal to insert on the dorsal tubercle of the humerus. By this method, a muscle lying below the wing can elevate the wing for flight and this keeps the mass of the bird stable. This muscle is used mainly for takeoff and is not necessary while in flight. It is particularly developed in species that hover and perform steep takeoffs (King & Custance 1982; King & McLelland 1984).
The tail acts as a rudder and brake. The tail feathers are attached to the pygostyle and radiate from it like a fan. Each feather can both move individually or as a whole, giving the bird fine control during flight. They can fan out to create drag and slow the bird down or twist to cause turning in flight (King & McLelland 1984; Raikow 1985).
CLINICAL NOTE
The pectoral muscles can be palpated to assess body condition and are also a site for intramuscular injections. However, pectoral injections should be avoided in birds relying on 100% flight efficiency: for example, birds of prey or wild birds due to be released.
Many birds (such as Galliformes) have fenestrated sternums, so when making intramuscular injections introduce the needle cranially, parallel to the sternum, to avoid puncturing the liver.
Propatagium
This triangular fold of skin extends from the shoulder to the carpus and forms the leading edge of the wing. It consists of thin dorsal and ventral layers of skin with a strong ligament running along the edge. This ligamentum propatagiale arises proximally on the deltopectoral crest and inserts onto the extensor process of the carpometacarpus. It has collagen at each end, is elastic in the middle and, when tensed, helps to achieve smooth airflow. Elastic branches fan out from the elastic part of the ligamentum and across the propatagium to insert on the dorsal antebrachial fascia of the distal ulna. In some large raptors a support ligament runs parallel to this tendon to give additional support when the wing is extended (Brown & Klemm 1990).
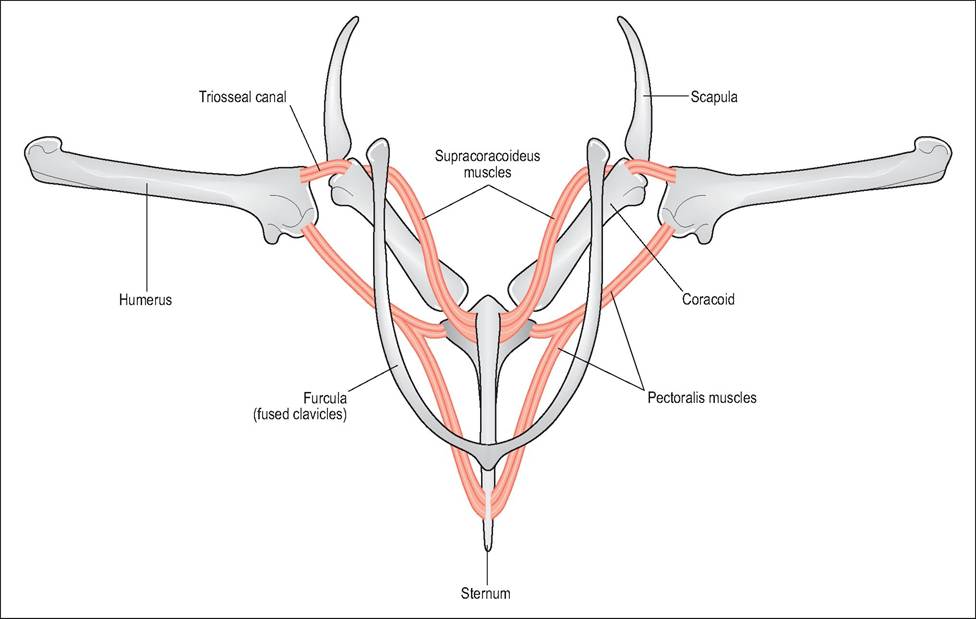
Figure 6.22 • Diagram of pectoral muscles demonstrating how both the supracoracoideus, which elevates the wing, and the pectoralis, which depresses the wing, have ventral origins. This keeps the heavy musculature close to the bird’s center of gravity.
CLINICAL NOTE
The propatagium, along with feathers, is essential for producing aerodynamic lift for flight. Propatagial tears or injury to the propatagial ligament are very difficult to repair and can render the bird flightless (Brown & Klemm 1990; Orosz 2002).
Pelvic limbs
The pelvic limb has been fused distally to absorb the shock of takeoff and landing (Figs. 6.23 and 6.24). It consists of femur, tibiotarsus, fibula, tarsometatarsus, metatarsal bone, and in most birds, four digits. It is covered by feathers and skin as far as the level of the intertarsal joint and is followed by scales below. In general the pelvic limb is not involved in flight and is used more for swimming, catching prey, and wading (Orosz et al. 1992) (Fig. 6.25).
Femur
This is a short bone that slopes cranially (almost horizontally) in order to ensure that the large feet lie under the bird's center of gravity. It articulates with the tibiotarsus and the
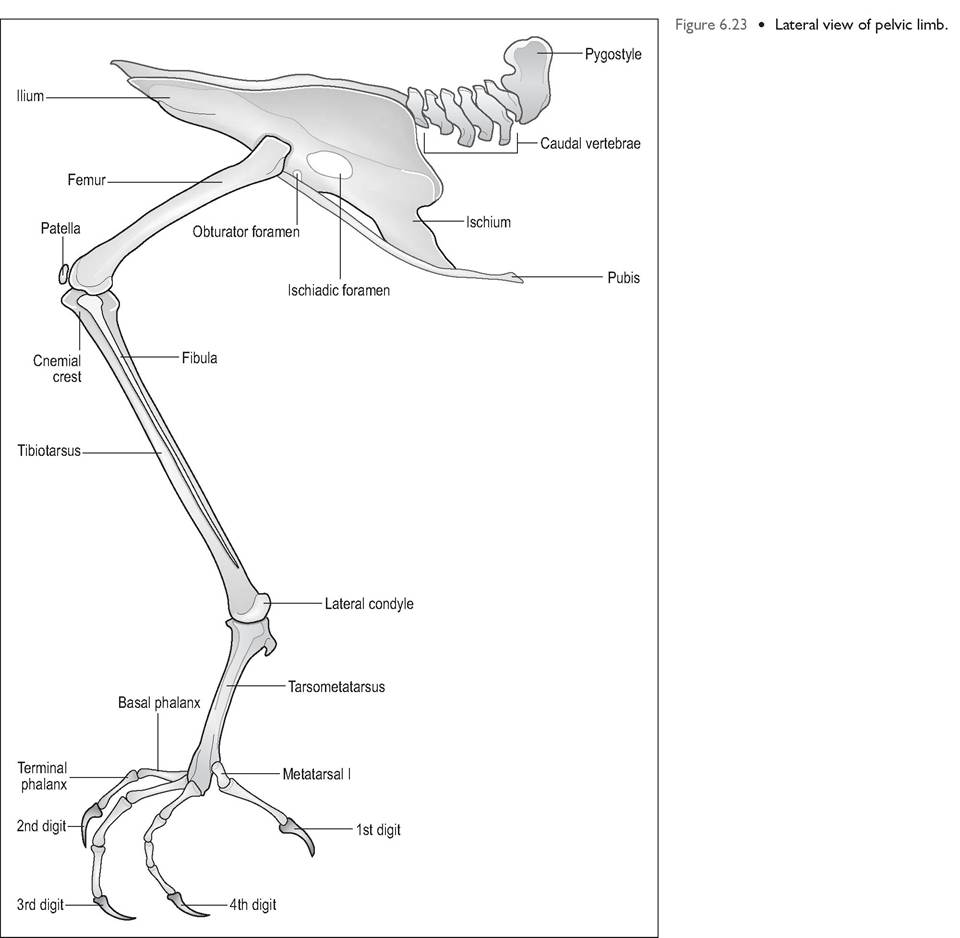
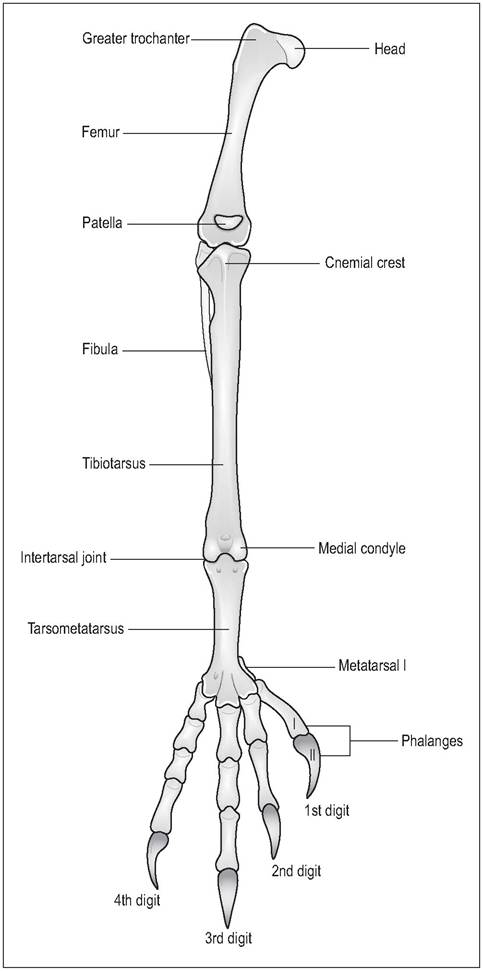
Figure 6.24 • Cranial view of right pelvic limb.
patella. As in mammals, the knee joint has two menisci, cranial and caudal ligaments, and collateral ligaments (Harcourt- Brown 1996). The femorotibialis muscle (the equivalent of the mammalian quadriceps) acts to extend the knee joint, and the patella is a sesamoid in its tendon of insertion. The main movements are flexion and extension.
Tibiotarsus
The tibiotarsus is the fusion of the tibia and proximal tarsal bones and is commonly called the drumstick in poultry. It is the largest bone on the pelvic limb (Evans 1996). A cranial crest known as the cnemial crest lies at the proximal tibia.
WHY DOES A DUCK WADDLE?
When birds walk they lean outwards to bring the center of gravity above the foot on the ground while the other leg is in the air. This is achieved by lateral rotation of the distal femur creating a waddling type gait. This is most noticeable in short-legged birds like ducks (Raikow 1985).
The fibula lies laterally and is short and not well developed in birds. The equivalent of the hock joint is between the tibiotarsus and the tarsometatarsus; movement here is mainly flexion and extension.
Tarsometatarsus
This is the fusion of tarsal bones 2, 3, and 4, to the corresponding metatarsal bones. The shape varies according to the locomotion of species. An elongated metatarsus conveys extra leverage for running or for takeoff in flight. The first metatarsus lies separate but is joined to the tarsometatarsus by ligaments. As it lies distally it can often be mistaken for a phalanx (Harcourt-Brown 1996; Raikow 1985; Smith & Smith 1997).
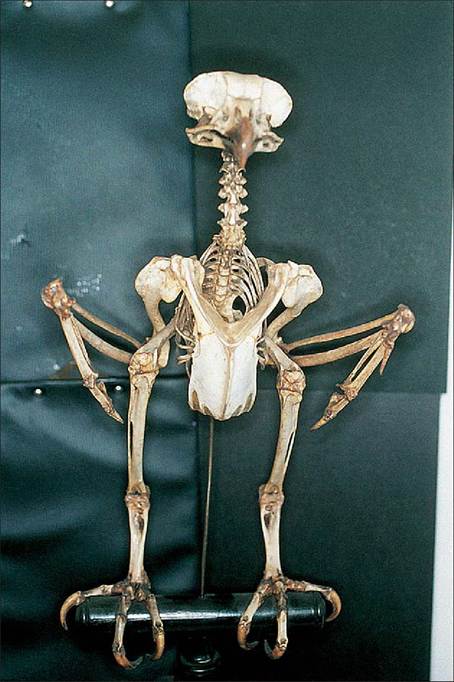
Figure 6.25 • Cranial view of skeleton of perching eagle owl (Bubo bubo).
Phalanges
Most birds have four digits with varying numbers of phalanges. Digit 1 will have two phalanges, digit 2 has three, digit 3 has four, etc. Digit 1 (the equivalent of our big toe) is often called the “hallux” and usually projects caudally. This is rudimentary in ground-dwelling birds and in fact the ostrich only has two toes (Bezuidenhout 1999; Evans 1996).
The main action of the digits is flexion and extension. When birds perch their tendons are flexed and small sprocketlike projections extend from these tendons and interdigitate to hold them in place. This reduces the amount of energy needed for perching.
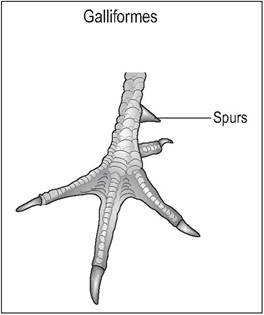
Figure 6.27 • Anisodactyl - Galliformes.
Galliforme foot showing spurs on the caudomedial aspect of the tarsometatarsus.
CLINICAL NOTE
112
The gastrocnemius tendon runs caudally along tibial cartilage to insert on the plantar aspect of tarsometatarsus. Calcium deficiency leading to nutritional osteodystrophy causes malformation of the tarsometatarsus. This causes the tendons to slip medially, causing lameness and rendering the bird unable to perch properly.
Arrangement of digits
Birds are called digitigrade because, unlike humans, they actually walk on their toes and not on their feet. The arrangement of the digits depends on their function. Feet can be used to perch, grasp prey, wade, or become fully webbed for swimming.
Anisodactyl
The majority of birds (Columbiformes, Galliformes, Passeriformes, Falconiformes) have an adaptation for perching or holding prey such that they have one toe facing backward and three facing forward (Fig. 6.26). Passerines also have an extra extensor muscle, the extensor hallucis longis to extend their first digit. Galliformes commonly have spurs on the caudomedial aspect of the tarsometatarsus (Fig. 6.27);
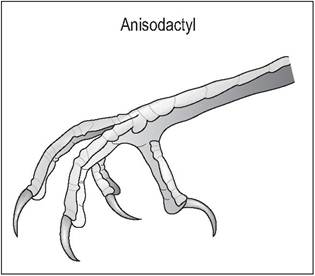
Figure 6.26 • Anisodactyl.
The majority of birds (e.g., passerines, pigeons) have one toe facing back and three facing forward. This is an adaptation for perching or holding prey.
these are more developed in the male (King & McLelland 1984).
Zygodactyl
Species like psittacines, have two (first and fourth) toes facing caudally and two (second and third) facing cranially. These species use their feet for grasping and climbing (Fig. 6.28). Owls and ospreys have a basic zygodactyl foot but can move the fourth digit from back to front (Raikow 1985).
Palmate
This is seen in ducks, geese, swans, and gulls and is when the cranial digits 2, 3, and 4 are webbed. The capacity to grip is sacrificed and the backward-pointing digit loses contact with the ground (Fig. 6.29).
KEY POINTS
• Thin bone cortices with internal medullary struts for strength
• Hens deposit calcium in medullary cavity prior to laying
• Rigid fused backbone for stability during flight - many species have weak link around T4
• Flight muscles all located ventrally on sternum, nearer to bird's center of gravity
• Neck and tail most flexible parts of backbone
• Pelvis incomplete ventrally, with fossae for kidneys
• Primary feathers insert on manus while secondary feathers insert on caudal ulna
Celomic cavities
Birds have no diaphragm and so have one body cavity called the celom that extends from the first thoracic vertebra to the vent. This cavity is not lined by serosa, as in mammals, but instead contains 16 separate cavities, 8 of which are air- sacs. The remaining eight cavities consist of three (two pleura and one pericardial) that resemble mammalian ones,
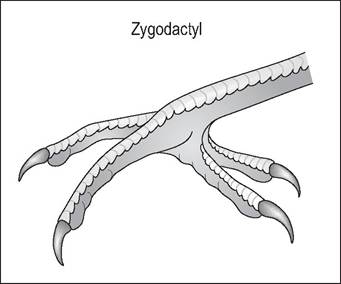
Figure 6.28 • Zygodactyl.
Psittacines have two toes facing caudally and two facing cranially. These feet are very dexterous and used for grasping food and climbing.
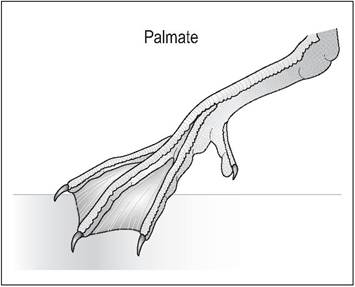
Figure 6.29 • Palmate.
Swimming birds like ducks, geese and gulls have webbing of the cranial digits II, III, and IV and are unable to grip.
and five peritoneal ones unique to birds. These cavities are of significance as they may influence the spread of pathology through the whole of the celomic cavity.
Pleural and pericardial cavities
The pericardial cavity is connected ventrally to the sternum and dorsally to the lung hilus and horizontal septum. The right and left pleural cavities are lined ventrally by a peritoneal sheet: the horizontal septum (Duncker 1979; King & McLelland 1975, 1984).
Peritoneal cavities
Birds lack an omentum, but a double-layered peritoneal sheet, the post-hepatic septum, stops the spread of disease by bringing in a healthy blood supply and divides the caudal cavity into three main parts. These are the middle intestinal cavity and two lateral hepatic cavities surrounding the liver. The hepatic cavities are in turn divided into the left and right ventral hepatic and left and right dorsal hepatic. The ventral hepatic ones are blind cavities between the ventral liver and body wall; the dorsal hepatic ones lie between the body wall and the dorsolateral liver. The intestinal cavity lies in the midline and contains the gastrointestinal tract, the gonads, and the oviduct, each of which lie suspended in their individual mesenteries (Duncker 1979; King & McLelland 1975, 1984).
CLINICAL NOTE
These cavities mean that egg peritonitis, for example, may be limited to the intestinal and left dorsal hepatic cavities only (King & McLelland 1984).