Cellularsenescence
Cellular senescence can be divided into two categories: replicative senescence and stress-induced premature senescence. Replicative senescence refers to the premature senescence that occurs after a limited number of divisions and a gradual shortening of telomeres at the ends of chromosomes.
This results in cell proliferation stagnation and loss of differentiation ability. Stress-induced premature senescence refers to the premature senescence that occurs in response to pathological stimuli, such as DNA damage and oxidative stress. These two types of senescence share many regulatory molecules, and they both cause cell cycle arrest mainly through the p53/p21 and p16Ink4a/retinoblas- toma protein signaling pathways. The number of senescent cells increases with age, obesity, and diabetes, and clearing senescent cells can alleviate many aging-related diseases and prolong the lifespan in mice. Senescent cells induce the formation of a complex, multicomponent SASP by secreting a range of cytokines, inflammatory factors, and adhesion factors. In the local microenvironment, the SASP alters the biological behavior of adjacent cells through autocrine and paracrine signaling. For instance, a systemic environment that shapes aging-related diseases has been reported to exist, as the infusion of young cerebrospinal fluid (CSF) improves the memory function of aged brains. This improvement is mainly mediated by serum response factor (SRF)-mediated oligodendrocyte progenitor cell (OPC) proliferation after exposure to young CSF. Elderly individuals often present with chronic low-grade inflammation, which is collectively referred to as immune aging. In the process of aging, the numbers of monocytes/macrophages, dendritic cells, and natural killer (NK) cells increase, possibly because of an increase in the number of aging cells, resulting in increases in the numbers of macrophages and NK cells, thereby eliminating aging cells. Immune cells may further induce the production of more pro-inflammatory cytokines and aggravate the progression of aging-related diseases.Senescence may be one of the main features of immune senescence. Premature T-cell failure may accelerate aging in multiple organs and systems, with thymic degeneration, mitochondrial dysfunction, genetic and epigenetic alterations, and imbalance in protein homeostasis being the four main hallmarks of T-cell senescence. In mice with mitochondrial transcription factor A (TFAM) deletion, T cells with mitochondrial dysfunction induce a variety of aging- related phenotypes, such as metabolic disorders, cognitive impairment, and cardiovascular diseases, which ultimately lead to the premature death of mice.
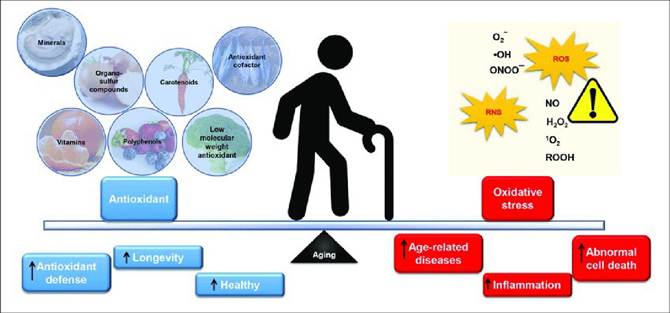
FIGURE 25.5 The balance of antioxidants and oxidative stress in aging
25.7.1 Mitochondrial Genome Damage
The mutation rate in mitochondria is 10-20 times faster than the nuclear DNA mutation rate. It is thought that mutations in mitochondria could 1. lead to defects in energy production, 2. lead to the production of ROS by faulty electron transport, and/or 3. induce apoptosis. Agedependent declines in mitochondrial function are seen in many animals, including humans (Boffoli et al., 1994). A recent report (Michikawa et al., 1999) shows that there are “hot spots” for age-related mutations in the mitochondrial genome, and that mitochondria with these mutations have a higher replication frequency than wild-type mitochondria. Thus, the mutants are able to outcompete the wild-type mitochondria and eventually dominate the cell and its progeny. Moreover, the mutations may not only allow more ROS to be made but may make the mitochondrial DNA more susceptible to ROS-mediated damage.
25.7.2 Telomere Shortening
Telomeres are repeated DNA sequences at the ends of chromosomes. They are not replicated by DNA polymerase, and they will shorten at each cell division unless maintained by telomerase.
Telomerase adds the telomere onto the chromosome at each cell division. Most mammalian somatic tissues lack telomerase, so it has been proposed that telomere shortening could be a “clock” that eventually prohibits the cells from dividing any more. When human fibroblasts are cultured, they can divide only a certain number of times, and their telomeres shorten. If these cells are made to express telomerase, they can continue dividing. However, there is no correlation between telomere length and the lifespan of an animal (humans have much shorter telomeres than mice), nor is there a correlation between human telomere length and a person’s age. Telomerase-deficient mice do not show profound aging defects, which we would expect if telomerase were the major factor in determining the rate of aging (Rudolph et al., 1999). It has been suggested that telomere-dependent inhibition of cell division might serve primarily as a defense against cancer rather than as a kind of “aging clock.”25.7.3 Hayflick Limit Theory
This theory of aging suggests that the human cell is limited in the number of times it can divide. The human cell's ability to divide is limited to approximately 50 times after which they simply stop dividing and hence die. Nutrition has an effect on cells with overfed cells dividing much faster than underfed cells. This theory indicates the need slow down the rate of cell division if we want to leave long lives.
25.7.4 Death Hormone Theory (DECO)
According to this we are born with roughly 12 billion of nerve cells and over a lifetime about 10% perish. This theory states that as we age the pituitary begins to release decreasing oxygen consumption hormone (DECO)or death hormone which inhibits the ability of cells to use thyroxin, a hormone produced by thyroid governing basal metabolism, the rate at which cell convert food to energy. The metabolic rate brings on and accelerates the process of aging.
25.7.5 Thymicstimulating Theory
The thymus is the master gland of the immune system.
The size of this gland reduces from 200 to 250 grams at birth to around 3 grams by age 60. Studies have shown that thymic factors are helpful in restoring the immune systems of children bone without them as well as rejuvenating the poorly functioning immune systems of elderly. Thymic hormones may also play a role in stimulating and controlling the production of neurotransmitters and brain and endocrine system hormones which means they may be the pacemakers of aging itself, as well as key regulators responsible for immunity.25.7.6 Caloric Restriction Theory
Underfeeding particularly restriction of calorie intake increases the life span. The effect of the restricted energy intake was to slow down the growth rate and to delay the attainment of maturity. In other words, the restricted calorie intake delayed the onset of senescence by prolonging the juvenile period.
25.7.7 The Rate of Living Theory
This theory states that we are each born with a limited amount of energy. If we use this energy slowly then our rate of aging is slowed.If the energy is consumed quickly aging is hastened. Other rate of living theories focuses on limiting factors such as amount of oxygen inhaled or number of heartbeats spent.
25.7.8 Order to Disorder Theory
From the time of conception to sexual maturation, our body undergoes a system of orderliness. Most of our energy is directed to fulfill a genetically determined plan for the orderly production and arrangement of an enormous number and variety of molecules. After sexual maturation, however, this energy starts to diminish in efficiency. Disorder occurs in molecules in turn causing other molecules to produce errors and so on. These chaotic changes in our cells, tissues and organs causes aging. Disorderliness varies from individual and this may be the reason why our tissues and organs deteriorate at different rates.
25.7.9 Psychosociological Theory
In addition to theories of aging based on molecules and cells, there also exists a “psychosociological” theory of aging.
As people grow older, their behavior changes, their social interactions change, and the activities in which they engage change. The psychosociological theory of aging can be divided roughly into four component theories: disengagement, activity, life-course, and continuity theories. Disengagement theory is based on hampered relationships between a person and other members of society. Activity theory emphasizes the importance of ongoing social activity and suggests that a person’s self-concept (self-perspective) is related to the roles held by that person. Life-course theory is based on the developmental stages proposed by German-born American psychoanalyst Erik H. Erikson. According to Erikson’s stages, maturity is a process that continues into old age, and in each stage the individual encounters new psychosocial demands. Continuity theory states that older adults try to preserve and maintain internal and external characteristics (e.g., values, personality, preferences, and behavior patterns) throughout life, despite changes in their health or life circumstances.25.8