OXIDATIVE DAMAGE
One major theory sees our metabolism as the cause of our aging. According to this theory, aging is a by-product of normal metabolism; no mutations are required. About 2-3% of the oxygen atoms taken up by the mitochondria are insufficiently reduced insufficiently to reactive oxygen species (ROS).
These ROS include the superoxide ion, the hydroxyl radical, and hydrogen peroxide. ROS can oxidize and damage cell membranes, proteins, and nucleic acids. Evidence for this theory includes the observation that Drosophila that overexpress enzymes that destroy ROS (catalase, which degrades peroxide, and superoxide dismutase) live 30-40% longer than controls (Parkes et al., 1998). Moreover, flies with mutations in the methuselah gene live 35% longer than wild-type flies. The methuselah mutants have enhanced resistance to paraquat, a poison that works by generating ROS within cells (Lin et al., 1998).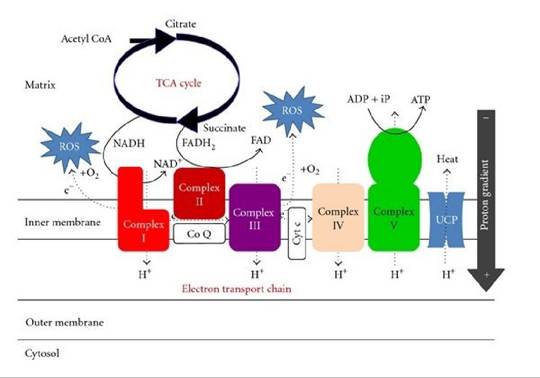
FIGURE 25.4 The electron transport chain responsible for ATP and ROS production in Mitochondria
These findings not only suggest that aging is under genetic control but also provide evidence for the role of ROS in the aging process. Individuals with mutations that increase the synthesis of ROS-degrading enzymes live much longer than wild-type. The evidence for ROS involvement in mammalian aging is not as clear. Mutations in mice that result in the lack of certain ROS-degrading enzymes do not cause premature aging. However, there may be more genetic redundancy in mammals than in invertebrates, and other genes may be up-regulated to produce related ROS- degrading enzymes. Migliaccio and colleagues (1999) have observed mutant mice that live one-third longer than their wild-type littermates. These mice lack a particular protein, p66. They develop normally, but the lack of p66 apparently gives them cellular resistance to ROS, and thus higher resistance to oxygen-induced stress on membranes and proteins. The p66 protein may be a component of a signal transduction pathway that leads to apoptosis upon oxygen stress, and it may be involved in mediating the life spans of mammals. Another type of evidence suggests that ROS may be important in mammalian aging: aging in mammals can be slowed by caloric restriction (Lee et al., 1999). However, caloric restriction can also have other effects, so it is not certain if it works by preventing ROS synthesis. Also, vitamins E and C are both ROS inhibitors, and vitamin E increases the longevity of flies and nematodes when it is added to their diet (Kakkar et al.,1996). However, results in mammals are not as easy to interpret, and there is no clear evidence that ROS inhibitors work as well as in invertebrates (Arking, 1998) (Figure 25.5).
25.7