Cerebellar function
The function of the cerebellum is to smooth and coordinate motor function for posture and movement. It does this by using subconscious proprioceptive information (see Chapter 6) to modulate motor activity of UMN nuclei and the motor cortex; these then influence LMN activity.
The cerebellum compares body position with desired motor output at the initiation, during, and at the termination of movement.Proprioceptive inputs to the cerebellum are primarily from somatic muscle spindles and Golgi tendon organs, and the vestibular apparatus. These inputs inform cerebellum about the position, status/tension and activity of the skeletal musculature, and the position and movement of the head. The cerebellum also receives copies of planned motor activity from the motor centres. With input about body and head position, and information about planned motor activity, the cerebellum can compare motor intention with action. It modulates motor output to achieve desired postures, and coordinated movement, involving a plethora of agonist and antagonist muscles. Ongoing proprioceptive feedback from the muscles and from the vestibular apparatus, permit constant comparison of the intended and achieved motor function. In mammals there are essentially no direct connections from the cerebellum onto LMN, thus the cerebellum cannot initiate movement. Consequently, in mammals of veterinary interest, cerebellar dysfunction does not cause paresis. However, birds have a prominent cerebellospinal tract and avian spinal cord LMNs depend more on cerebellar than cortical control. While cerebellar dysfunction does not cause paresis in domestic mammals, it does disrupt posture (static and dynamic), and the spatial accuracy and temporal coordination of movement.
Cerebellar connections
Cerebellar afferents convey proprioceptive information, or information relevant to the planning and execution of motor activity.
1. Proprioceptive input from limbs, body and head is represented ipsilaterally and informs the cerebellum about the distribution/location of body parts and status of muscles and joints. Afferent fibres include the spinocerebellar, vestibulocerebellar and tectocerebellar pathways.
2. Input about planned motor activity. The cerebellum needs this to establish the appropriate postural platform before the activity is initiated and, then, to regulate and coordinate muscle activity throughout that motor action.
(a) Input relevant to the coordination of functions originating from the motor cortex (learned, complex voluntary movement) is via the corticopontocerebellar pathway to the contralateral cerebellar hemispheres. There is a direct correlation between the extent of development of this system and the level of development of a species’ motor skills. Thus, it is far better developed in primates and human, which have highly developed control of digits, facial muscles (facial expression), tongue and laryngeal musculature for speech. The corticopontocerebellar pathway from the cerebral cortex travels through the internal capsule and the crus cerebri to synapse in nuclei in the pons. Post-synaptic fibres decussate and ascend via the middle cerebellar peduncle into the cerebellum. The return pathway is from the cerebellar cortex to the cerebellar nuclei (e.g. lateral nucleus). Post-synaptic fibres decussate and exit the cerebellum via the rostral cerebellar peduncle, travel rostrally to the synapse in the thalamus and then to the cerebral cortex.
(b) Input relevant to coordination of the extrapyramidal function (semiautomatic movement, posture and locomotion) originates from the cerebrum, thalamus and midbrain, and projects to the olivary nuclei in the medulla oblongata. These nuclei project to the contralateral cerebellum. The cerebellum makes return connections via the deep cerebellar nuclei to the contralateral extrapyramidal nuclei of the forebrain (cortex and basal nuclei) and brainstem.
Efferent fibres from the cerebellar cortex are from the Purkinje cells. They are inhibitory and the majority synapse on the deep cerebellar nuclei, inhibiting them. The output from the deep cerebellar nuclei forms the majority of efferent fibres from the cerebellum; it is facilitatory to motor systems, thus loss of cerebellar cortical output usually results in excess UMN activity and spasticity (Figs. 7.7, 7.8). Note: output from the vestibulocerebellum (flocculonodular lobe) bypasses the deep cerebellar nuclei and synapses directly onto the vestibular nuclei.
Summary of cerebellar connections
■ All afferent fibres to the cerebellar cortex are excitatory.
■ All efferent fibres from the cerebellar cortex (Purkinje cells) are inhibitory.
■ Purkinje cells are directly excited by cortical afferents but are also inhibited by interneurons. The output of Purkinje cells depends upon the balance between excitation and inhibition.
■ Deep cerebellar nuclei neurons are all excitatory to nuclei of pyramidal and extrapyramidal systems.
■ Deep cerebellar nuclei are inhibited by Purkinje cells and excited by afferent collaterals of afferent fibres.
■ Deep cerebellar nuclei can be excitatory or silent to motor nuclei, but never inhibitory.
Role of cerebellum in posture, locomotion and movement: Setting the postural platform
Posture is the position of the body or body parts. It results from coordinated activity of extensor and flexor muscles acting around numerous joints of the limbs and vertebral column.
The cerebellum is the centre that coordinates activity in agonist/antagonist muscles acting around the joints throughout the body. But it also has a critical role in establishing the postural platform on which movement is superimposed. For example, a cat cannot catch an object with one paw without first redistributing its weight over the other three legs. Setting the body up to support the movement is called setting the postural platform.
To set the appropriate tone in the postural muscles, the cerebellum must be informed about what movements are being planned by the executive and senior management motor systems. It must also know exactly where the body parts are at that moment and what they are doing with respect to supporting posture. Thus it compares input from the motor planning centres with subconscious proprioceptive input.
The cerebellum then influences output of the UMN to the LMNs controlling postural muscles, to set the postural platform - this may involve redistribution of the weight. Having coordinated that postural adjustment, the cerebellum receives the new proprioceptive input from the activated postural muscles. In turn, the cerebellum feeds back to the motor management systems informing them that the postural platform has been set and the motor activity can go ahead.
If the postural platform is not established, then the movement cannot occur. This is called postural paralysis (Fig. 7.9).
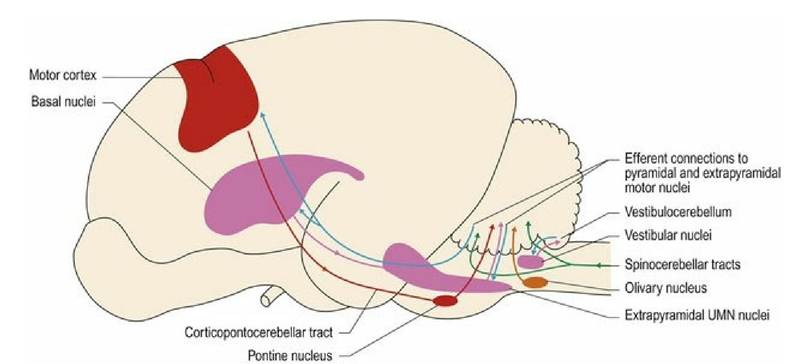
Fig. 7.8 Cerebellar connections. The cerebellar efferent fibres (blue) may have a direct inhibitory effect
(e.g. on vestibular nuclei), or have a facilitatory effect on UMN centres; but this effect is inhibited by
Purkinje cells of the cerebellar cortex.

Fig. 7.9 Setting the postural platform. Barney has flexed his left thoracic limb. Before he could do that, he had set the postural platform to permit the movement. He had to unload his body weight from the left foreleg and redistribute the weight onto the right thoracic limb, which is in obvious extension. Barney has also shifted his weight caudally onto his seated hindquarters. Additionally, visual input has resulted in a change in the head position, tilting it upwards. This will stimulate the vestibular system (see Chapter 8) and the neck proprioceptors; vestibular stimulation will further facilitate the extension of the thoracic limb.
Thus, an apparently simple movement (flexing one limb) actually requires coordination of most of the muscles in the body, limbs and neck.Cerebellar dysfunction
In summary, three different syndromes may occur with lesions in different parts of the cerebellum (Fig. 7.4A). If the cerebellum is diffusely affected, then a combination of signs may occur.
1. Vestibulocerebellar signs. The animal has a disturbance of equilibrium with swaying posture, wide-based stance and falling to the side when moving around. It may also have nystagmus and strabismus, but spasticity and tremor are not apparent.
2. Spinocerebellar signs. Hypermetria and hypertonus (spasticity) result in exaggeration of spinal reflexes, gait and postural responses such as hopping, wheelbarrowing, hemiwalking, etc.
3. Pontocerebellar signs. These affect feedback pathways between the cerebellum and the forebrain and can result in asynergia in which there is loss of harmony and synchrony in movements. Signs include dysmetria and overshooting of body parts, such as the head and limbs, and tremor that is exacerbated as the animal attempts to make a voluntary movement; this is known as an ‘intention tremor’.
Cerebellar lesions can result in either or both of the following:
■ Inadequate processing of incoming proprioceptive information resulting in subconscious proprioceptive deficits
■ Inadequate output for modulating motor activity, thus motor activity is usually excessive.
Consequently, the following signs may be seen.
Ataxia is the uncoordinated or inconsistent movement of the body and limbs. This is caused by reduced/no processing of proprioceptive input to the cerebellum. Thus the cerebellum does not know the position and status of body parts and cannot coordinate postural and locomotory muscles.
Dysmetria is abnormal rate, range or force of movement. Failure to receive incoming subconscious proprioceptive information results in incorrect postural adjustment and there will be delays in the initiation and termination of movements.
Similarly, if the cerebellum cannot continually monitor subconscious proprioceptive input during movement, there will be failure to regulate the rate, range and force of movement. In the case of cerebellar lesions this often results in hypermetria with increased rate, range and force of movement. Hypermetria is characterised by overshooting of body parts during movement, and exaggerated stepping action (goose-stepping) during locomotion in animals. As the diseased cerebellum cannot receive subconscious proprioceptive feedback informing it that adequate movement has occurred, it fails to facilitate termination of movement (Fig. 7.10).Fig. 7.10

Hypermetric thoracic limb gait in a young Arabian horse with cerebellar degeneration
(courtesy of Prof. Joe Mayhew, IVABS, Massey University).
Spasticity occurs because of inadequate inhibition of UMNs. This leads to excessive muscle tone (extensor dominance) and also contributes to hypermetria. Note that paresis is NOT a feature of cerebellar disease. The rostral lobe of cerebellum particularly, exerts an inhibitory effect on antigravity muscles. Thus lesions of the rostral lobe can result in marked extensor activity including opisthotonus and thoracic limb extension.
Tremor arises due to failure to coordinate activity of the contracting agonist and relaxing antagonist muscles acting around a joint. Thus the muscle groups fight each other causing the joint to oscillate between flexion and extension. The classic presentation is known as an ‘intention tremor’ in which a tremor occurs associated with goal-directed movements such as an animal reaching forward with its head.
Vestibular signs occur due to involvement of the vestibulocerebellar connections or the vestibulocerebellum. The vestibulocerebellum receives proprioceptive input about head proprioception from the vestibular system. Inadequate vestibulocerebellar function results in reduced output from the cortex of the vestibulocerebellum. As this output is from inhibitory Purkinje cells to the vestibular nuclei, there is usually excessive activity of the vestibular nuclei in the brainstem. This occurs with lesions in the flocculonodular lobe or in the caudal cerebellar peduncles that convey connections between the vestibular nuclei and cerebellum. Therefore on the side with the lesion, there will be increased activity in the vestibular nuclei, resulting in increased ipsilateral extensor (anti-gravity) muscle tone and a head tilt to the side opposite to the lesion. This results in the paradoxical vestibular syndrome, which is explained more fully in Chapter 8.
Menace response deficits can occur in cerebellar disease in which, despite normal vision, the animal fails to blink in response to a threatening stimulus. The pathway by which this occurs is unclear but may be due to the visual pathway connecting to CN VII nuclei for blinking, via the cerebellum (corticopontocerebellar tract) or it could be due to failure of cerebellar facilitation to forebrain motor cortex.
Mostly signs of cerebellar dysfunction are ipsilateral, except in paradoxical vestibular syndrome in which the signs suggest a contralateral lesion.