COMMUNICATIONS OF STIMULUS BY THE GUSTATORY SYSTEM
Saliva carries chemical substances that enter taste buds through pores at their top, where they bind to receptors located on the membrane of microvilli within taste cells. This binding triggers depolarization of the taste cell membrane.
The specific mechanisms of membrane depolarization vary depending on the type of taste molecules binding to their respective receptors. For instance, salty taste transduction involves the influx of Na+ ions through amiloride-sensitive Na+ channels (ENaC), leading to membrane depolarization (Figure 13.1). However, the exact transduction pathways for salty tastes remain incompletely understood. One hypothesis suggests that the transduction process may involve the opening of voltage-gated Ca2+ channels, resulting in the influx of Ca2+ ions and subsequent release of neurotransmitters. Notably, taste cells responsible for detecting salty tastes lack well-defined synapses, and the neurotransmitter involved in their signaling remains unidentified. Additional research is needed to further elucidate the intricate mechanisms underlying salty taste perception.Elevated intracellular Ca2+ levels, combined with membrane depolarization, trigger the release of ATP via gap junction channels in the plasma membrane. Released ATP acts on sensory nerve endings, initiating generator
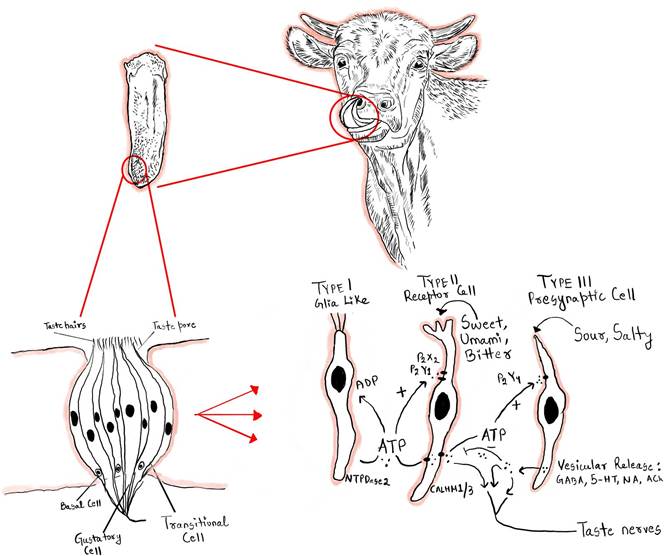
FIGURE 13.1 The gustatory organ of cattle for taste sensation is located in the tongue.
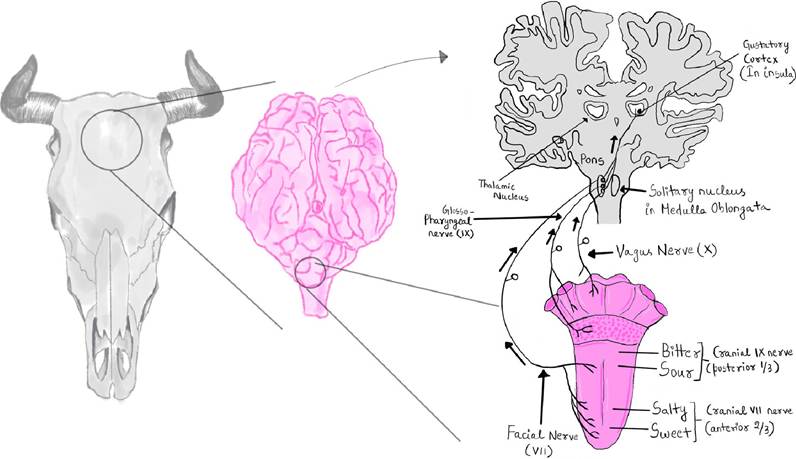
FIGURE 13.2 Central control of nerves for gustatory physiology of cattle.
potentials. If these generator potentials reach the threshold potential, action potentials are generated. Notably, taste cells responsible for sweet taste detection lack well-defined synapses, adding complexity to the transmission of signals from taste buds to sensory nerves. Additional research is needed to fully elucidate the intricate mechanisms involved in sweet taste transduction. Sweet taste transduction involves G protein-coupled receptors that activate phospholipase C (PLC). Upon activation, PLC generates two intracellular second messengers, inositol trisphosphate (IP3) and diacylglycerol (DAG), by hydrolyzing phosphatidylinositol 4,5-bisphosphate (PIP2). IP3 binds to calcium channels on the endoplasmic reticulum, causing them to open and allow Ca2+ to flow into the cytosol. There is speculation that taste-selective cation channels (TrpM5) are sensitive to changes in calcium levels, with IP3 playing a crucial role in their activation. This activation leads to the influx of Na+ ions through TrpM5, resulting in the generation of a depolarizing receptor potential in taste receptor cells (Figure 13.1).
13.4