Development of the Male Reproductive Organs
The early stages of morphologic differentiation of the reproductive organs follow a pattern common to the two sexes. In both, the gonadal primordium appears as a thickening of the celomic epithelium on the medial aspect of the mesonephros.
It projects as a swelling when the underlying mesenchyme proliferates (Fig. 5.7A/5). Cords of cells that develop from the covering epithelium penetrate the interior of the swelling (Fig. 5.7B/5). These cords shortly incorporate the primordial germ cells, which rather surprisingly have a distant origin in the endoderm of a restricted portion of the yolk sac, where they are identifiable by their large size. They reach the gonad by migration over the gut and its mesentery, but carriage in the bloodstream also seems possible.An early indication that the gonad will become a testis is provided by a marked mesenchymal condensation (tunica albuginea) below the celomic epithelium. Now isolated from the surface epithelium, the cords increase in size and in complexity of arrangement (Fig. 5.8/3). They connect to a plexus or network (rete) within the testis. On the other side the plexus makes contact with the blind ends of the few tubules that have survived the general regression of the mesonephros (Fig. 5.8B/3-5). Differentiation within the cell cords permits recognition of two cell lineages. One provides the sustentacular (Sertoli) cells of the seminiferous tubules. The second, contributed by the primordial germ cells, provides the germinal epithelium. During fetal development the primordial germ cells differentiate into gonocytes, which after birth give rise to spermatogonia. At puberty, the spermatogonia proliferate and differentiate to supply cells that undergo meiosis and spermiogenesis to form male gametes (see Fig. 5.39). Sections through the adult testis show seminiferous tubules cut in various planes.
The walls of the highly convoluted tubules are lined by a stratified germinal epithelium consisting of cells in various stages of differentiation. Supporting Sertoli cells nourish the germ cells. The Leydig cells produce the steroid testosterone that is essential if spermatogenesis is to continue. Their progenitors, like those of Sertoli and primordial germ cells, presumably migrate from the mesonephros during fetal development to become embedded in a mesenchymal interstitium, and around puberty, when the process of spermatogenesis is initiated, asecond generation of Leydig cells develops.
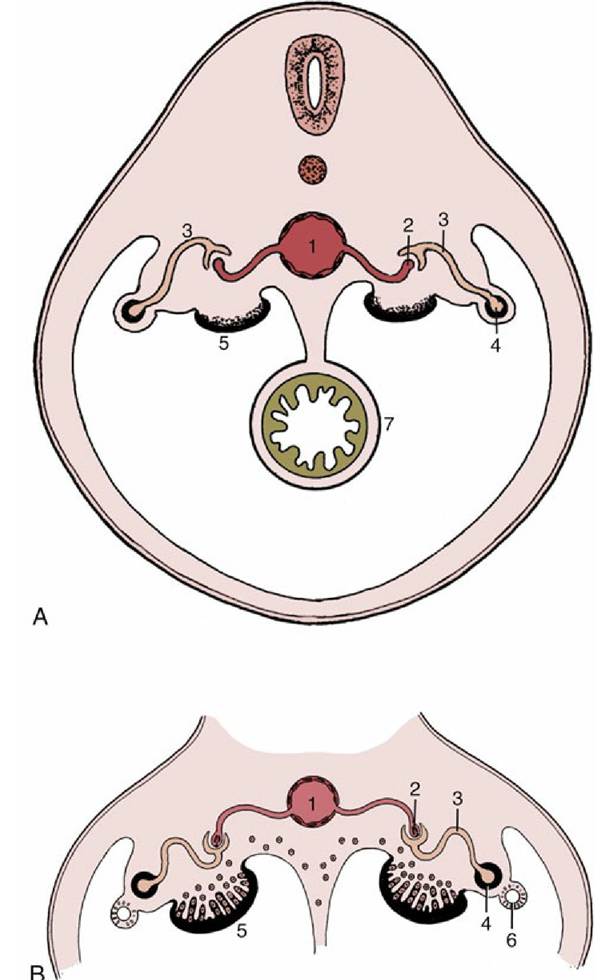
FIG. 5.7 (A) Early development of the indifferent gonad. (B) Invasion of the gonad by epithelial cords, which then incorporate primordial germ cells. 1, Aorta; 2, capillary tuft (in nephron); 3, nephron (tubule); 4, mesonephric duct; 5, gonad; 6, paramesonephric duct; 7, gut.
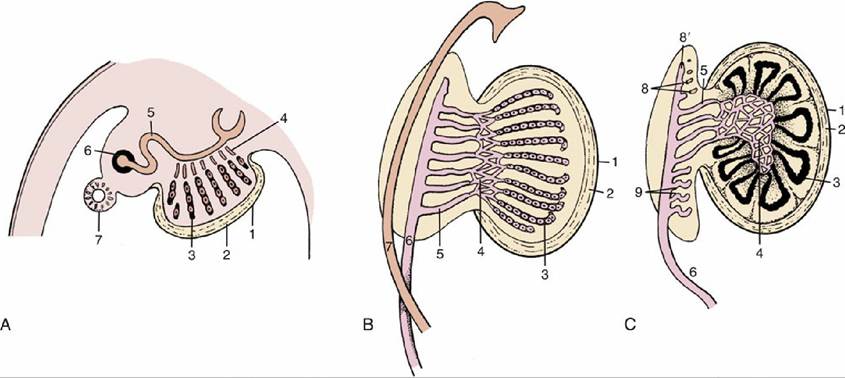
FIG. 5.8 Three stages in the development of the testis. (A) The epithelial cords are isolated from the surface epithelium by the formation of the tunica albuginea. (B) The epithelial cords, rete, and mesonephric tubules have interconnected. (C) The epithelial cords become seminiferous tubules, and the mesonephros is gradually transformed into part of the epididymis. 1, Celomic epithelium; 2, tunica albuginea; 3, epithelial cords, seminiferous tubules; 4, rete testis; 5, mesonephric tubules, efferent ductules; 6, mesonephric (later deferent) duct; 7, paramesonephric duct; 8, cranial remnant of mesonephric tubules (aberrant ductules); 8', remnant of 6 (appendix of epididymis); 9, caudal remnant (paradidymis).
The initial formation of the seminiferous cords is followed in later fetal life by canalization of the cords to create a series of passages leading to the mesonephric duct, which thus becomes the outlet for the gamete products of the testis.
The peripheral parts of the cords become seminiferous tubules, the central parts become the rete testis, and the mesonephric tubules become the efferent ductules (Fig. 5.8C). The first part of the mesonephric duct convolutes and forms the duct of the epididymis within the dense connective tissue of that organ; the remaining part retains a straighter course, and as the deferent duct (Fig. 5.5/3), it opens into that part of the cloaca that becomes the urogenital sinus (Fig. 5.5/6). Glandular proliferation of the lining of the duct toward its termination produces the ampullary thickening, whereas in most species but not in carnivores, a subterminal budding enlarges as the vesicular gland (Fig. 5.9/5). In some species a final short passage, the ejaculatory duct, persists, but in others later adjustments cause the deferent and vesicular ducts to open separately. Gonadal enlargement causes the testis to hang within a fold (mesorchium) arising from the regressing mesonephros. The duct is carried within this supporting fold, which in its caudal stretch inclines medially to form with its neighbor the genital fold of peritoneum that helps subdivide the peritoneal cavity of the pelvis. The testis later migrates outside the abdomen (p. 163) before the initiation of spermatogenesis.The division of the cloaca has been described (p. 136). The caudal part of the sinus constitutes the pelvic part of the urethra. Outgrowths from its lining differentiate into the prostate and bulbourethral glands in a species-characteristic fashion (Fig. 5.9). The greater part of the male urethra lies within the penis and has a different origin. Thickenings appear around the margin of the urogenital membrane in the indifferent stage (Fig. 5.10). One thickening, ventral and median, constitutes the genital (phallic) tubercle or swelling (Fig. 5.10/1), which gives rise to the greater part of the penis; other thickenings that are more lateral in position contribute the scrotum. A further urogenital fold that appears medial to each scrotal swelling makes an additional contribution to the penis.
A groove extends along the (initially) dorsal surface of the genital tubercle; it is gradually closed by the approach and merging of these urogenital folds. This process is rather complex because the lining of the penile urethra is provided by an extension of the endoderm of the urogenital sinus, although the initial swellings have ectodermal coverings. The corpus spongiosum (spongy body) of the penile urethra directly continues the bulbar tissue of the pelvic urethra, while the corpus cavernosum penis forms within the genital swelling. The lateral swellings grow and join together to form the scrotum, which retains evidence of its bilateral origin in a median raphe and septum.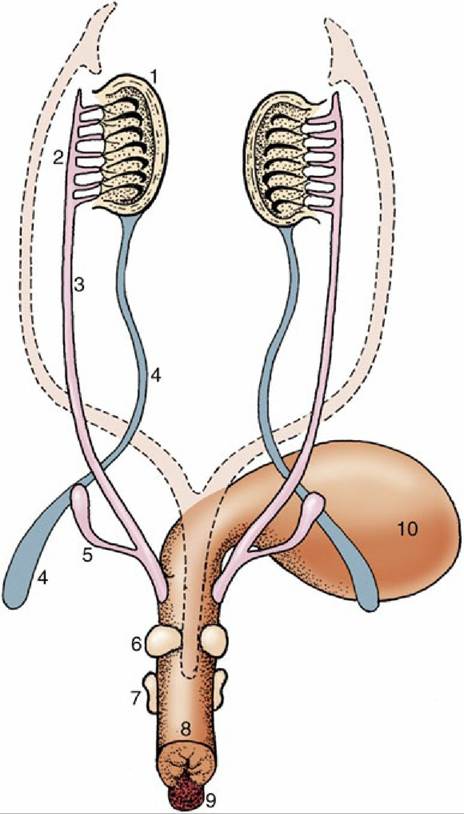
FIG. 5.9 Differentiation of the urogenital sinus. Note the budding of the prostate and bulbourethral glands and the enlargement of the genital tubercle. The regressed paramesonephric ducts are indicated by the broken lines. 1, Testis; 2, epididymis; 3, deferent duct; 4, gubernaculum; 5, vesicular gland; 6, prostate; 7, bulbourethral gland; 8, urogenital sinus (urethra); 9, genital tubercle; 10, bladder.
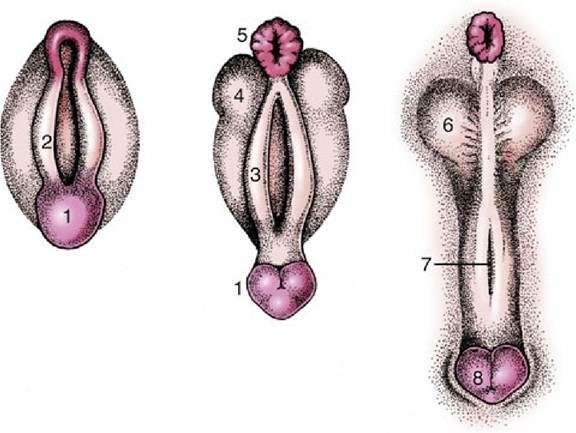
FIG. 5.10 Development of the male external genitalia. 1, Genital tubercle; 2, cloacal fold; 3, urogenital fold; 4, lateral (scrotal) swelling; 5, anus; 6, scrotum; 7, groove closing to form the penile urethra; 8, glans penis.
Differentiation of the male efferent duct system, accessory glands, and external genitalia depends on the presence of testosterone, the male sex hormone produced by the developing testes. The testes also produce several other hormones—for example, the antimullerian hormone (AMH) and insulinlike factor 3 (descendine), respectively responsible for the disappearance of the mullerian duct and the outgrowth of the gubernaculum, respectively. Without exposure to these three hormones the genital tract would develop in the female direction. Removal of the pituitary by decapitation in the fetal period does not disturb the production of these hormones by the testis (Fig. 5.11A and B).