Differentiation of Other Tissues
Mesodermal cells on each side of the notochord condense into a series of block-like paired masses. These are the segmentally arranged somites, which in turn develop into vertebrae and muscles.
Mesoderm lateral to the somites is called nephrogenic (intermediate) mesoderm. It will give rise to urogenital organs (Fig. 3-4).The most lateral mesoderm, the lateral plate mesoderm, splits into two layers, forming a cavity, the coelom. The coelom will eventually become the thoracic, abdominal, and pelvic cavities.
The outer layer of the lateral plate mesoderm and the adjacent ectoderm make up the somato- pleure, which forms part of the body wall and enters into the formation of the fetal membranes. The inner layer of the lateral plate mesoderm and the endoderm form the splanch- nopleure, which forms the wall of the gut (Fig. 3-5).
Differentiation of the relatively indifferent cells of each of the three germ layers to form specialized tissue cells is called histogenesis. Much is known about when and where various tissues and organs develop, and developmental biologists are beginning to unravel the
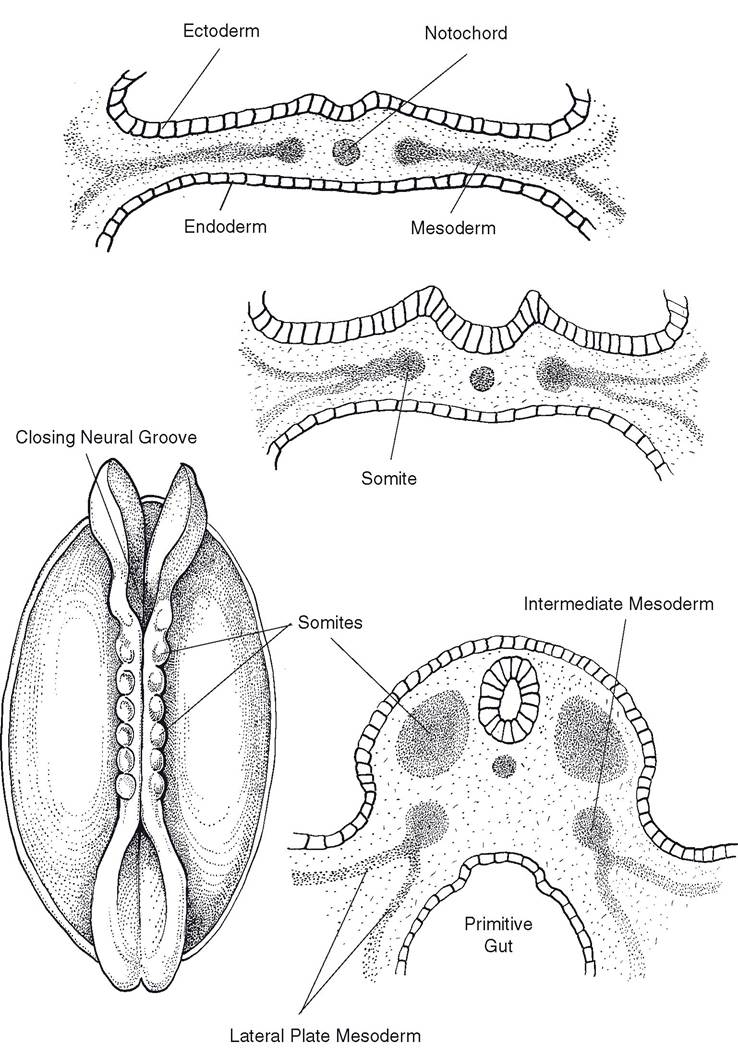
Figure 3-4. Differentiation of mesoderm. Somites form most medially. These will give rise to the muscles and bones associated with the vertebral column. Lateral to the somites is the intermediate mesoderm, which will differentiate into tissues of the urogenital system. Most laterally, the lateral plate mesoderm will become part of the gastrointestinal wall and body wall.
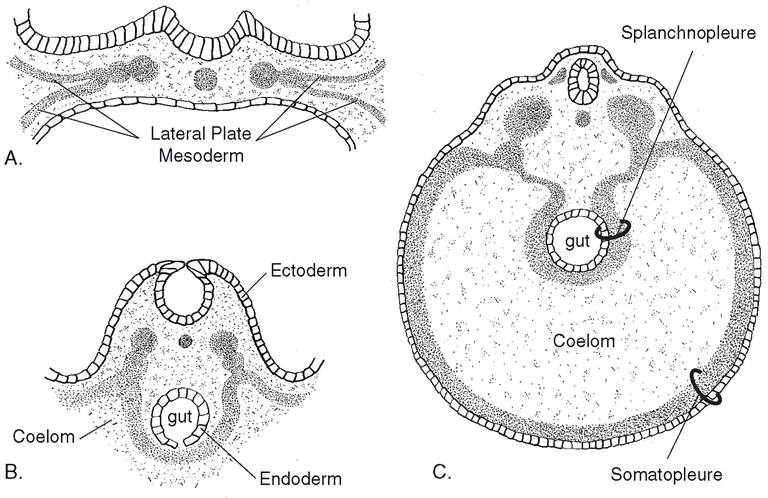
Figure 3-5. A) Lateral plate mesoderm splits early in development, creating the coelom (early body cavity). B) The external layer associates with overlying ectoderm, while the internal layer associates with endoderm of the developing gut.
C) Ectoderm plus the external layer of lateral plate mesoderm constitutes the somatopleure; it will become the body wall. The internal mesoderm and endoderm are called splanchnopleure, which becomes the wall of the gastrointestinal tract.Table 3-1. The Germ Layer Origin of Tissues

molecular and genetic events that underlie development. For instance, cell surface molecules that instruct cells to migrate and aggregate have been identified, as have others that cause cells to change the expression of their genes. once a cell has altered the expression of its genome so as to assume a more specialized role, it is said to be committed. Each of the three germ lines is committed to form certain kinds of tissues (Table 3-1). Differentiation
occurs as these cells assume the appearance and functions characteristic of the cell type to which they are committed.
Embryonic stem cells are capable of differentiating into multiple tissue types, a characteristic called pluripotency. Unlike most differentiated tissues, they can survive in long-term culture (outside the body). Embryonic stem cells are profoundly interesting to biomedical researchers, as it appears that cultured stem cells can be induced to differentiate into a wide variety of tissue types. This technique offers promise of a novel way of treating diseases involving loss of normal tissue. A few of the many possibilities include using stem cells to create (1) hematopoietic cells to regenerate normal blood precursors for treatment of blood and immune disorders, (2) cartilage and bone to replace damaged skeletal components, and (3) neuronal precursors to replace the neurons lost in Parkinson’s disease or Alzheimer’s disease. Research with human embryonic stem cells is fraught with contentious ethical considerations, since these cells are generally acquired from aborted embryos and fetuses. Research using animal embryos tends to be less ethically problematic, but it will naturally lead to intense debate as techniques developed in animal models are extrapolated for use in human medicine.