DIGESTIVE SYSTEM
The good news is that, because the trachea has rigid rings and the carotid arteries are protected by the cervical vertebrae, it would be difficult to strangle a bird. The bad news is that birds can easily asphyxiate by excessive pressure on their sternum and abdominal muscles, so be careful when handling them (King & Payne 1964).
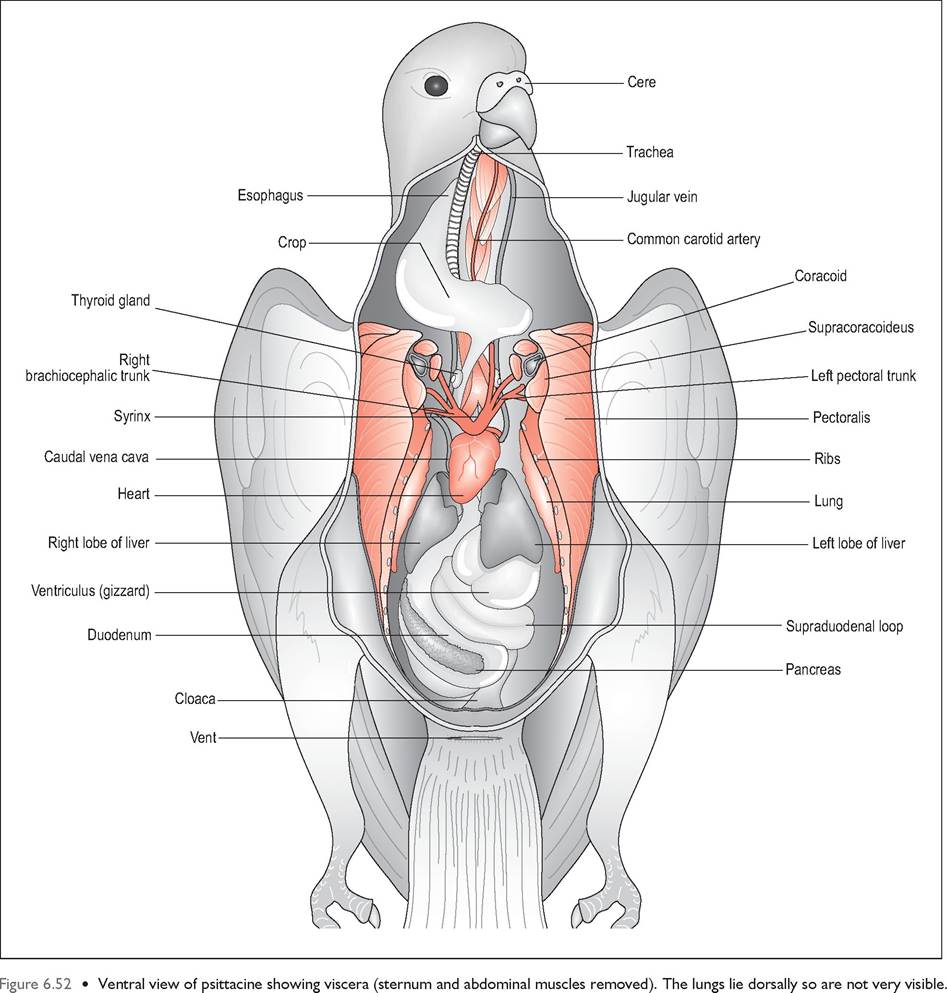
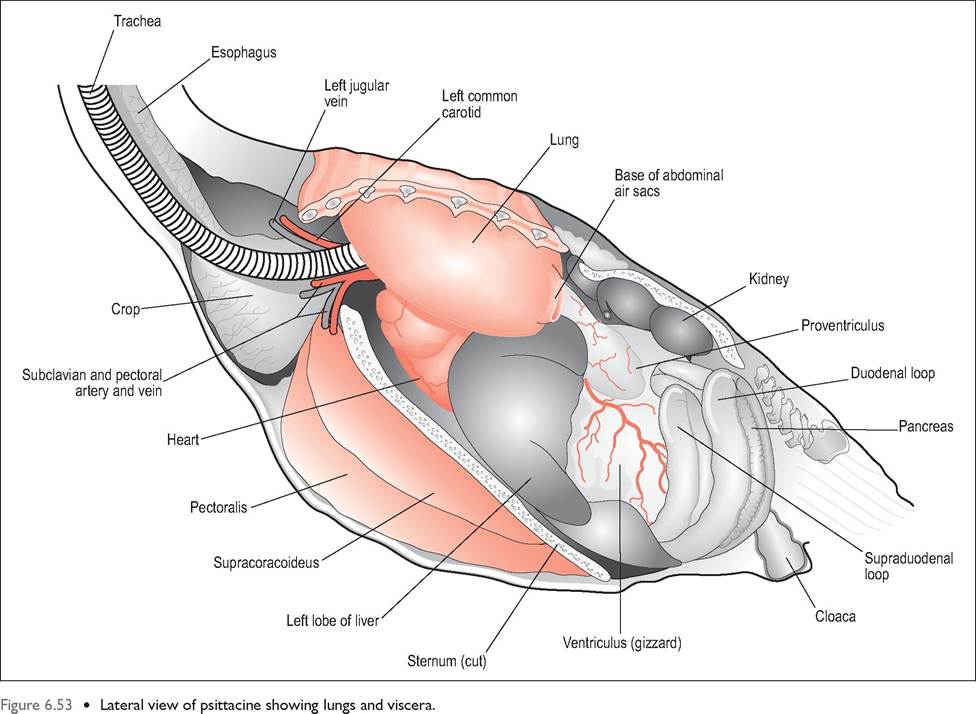
The avian gastrointestinal tract (Figs. 6.52 and 6.53) is relatively short, with low volume to keep the bird lightweight for flight. Consequently, birds ingest small amounts frequently and extract energy and nutrients rapidly to sustain their high metabolic rate. Transit times ranging from as little as 16 minutes to 2 hours are found in passerines. Birds also have
CLINICAL NOTE
Macrophages in the parabronchial epithelium filter out a lot of potential pathogens. The cranial airsacs are consequently filtered more than the caudal airsacs and this means a disease like aspergillosis is much more common in the caudal airsacs (Fedde 1993) (Fig. 6.47).
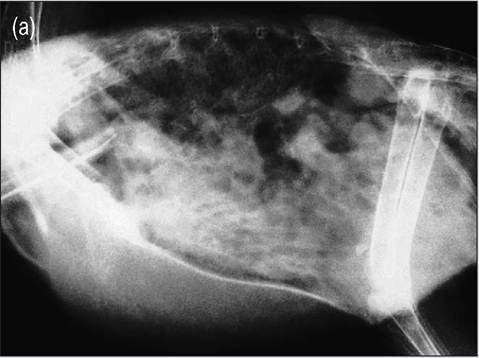
Figure 6.47 • Radiograph of an African gray parrot with chronic aspergillosis of thoracic and abdominal airsacs.
(a) Lateral view
(b) Ventrodorsal view
an extremely efficient digestive system, passing remarkably small amounts of excreta in contrast to the amount of food eaten.
Birds have no teeth so no time is spent on chewing and food passes rapidly to the crop for storage and into the gizzard for mechanical digestion. This heavy organ located at the bird's center of gravity has taken on the role of mammalian molars in grinding down the unmasticated food. Herbivorous birds like the ostrich and chicken also have well-developed ceca for food breakdown.
Beak
The beak (synonym bill) replaces the lips and teeth of mammals and consists of bone, vascular dermis and a
CLINICAL NOTE
Birds have low fat reserves due to the negative effects of excess weight. Small birds like budgies and canaries should not be fasted prior to anesthesia as they have low glycogen reserves and rapidly become hypoglycemic.
modified heavily keratinized, germinal layer. The keratin layer of the beak is called the rhamphotheca, with the rhinotheca covering the upper beak and the gnathotheca covering the lower beak. The lower beak slips inside the upper beak, fitting into a notch on the roof when the jaws are closed. Soft folds of keratin at the angles of the mouth form the rictus.
The epithelium has a very thick stratum corneum that contains hydroxyapatite, calcium, phosphate, and keratin and gives the beak its hardness. The vascular dermis is extremely thin and is sandwiched between the horny epidermis above and the periosteum of the premaxilla and mandible below.
Beak keratin appears in two forms. The weight bearing horn (tomia) found at the cutting edges and tips of both beaks performs the function of teeth in cutting up food. A second covering horn lies externally on the non-contact surface of the beak. Newly hatched chicks have a horny pro-
BEAK & TONGUE MODIFICATIONS IN WATERFOWL
Many waterfowl have modified their beak and tongue into a specialized filter system for sifting out food particles from water. Ducks, geese, and swans have a thick, fleshy tongue with the rostral border modified into a scoop with bristles on the lateral margins. There is a shallow median groove on the dorsal tongue and a flesh caudal eminence, the torus linguea. The upper and lower beak have lamellae into which the bristles fit (Fig. 6.48). Water is sucked in to fill the dorsal groove in the tongue and this is then pressed against the palate and filtered laterally by the bristles, which retain the food particles. In some species the bill tip contains sensory organs called Herbst corpuscles in the dermis to help discriminate food particles while feeding (McLelland 1979).
Flamingos have a specialized beak that filters organisms from the mud or water. At hatching the bill is straight but develops a bend when the beak matures. The bird feeds with the bill inverted so that the mandible is uppermost. The piston-like tongue moves food and water in and the solid particles are filtered out by the lamellae of the upper and lower beak (Fig. 6.49).
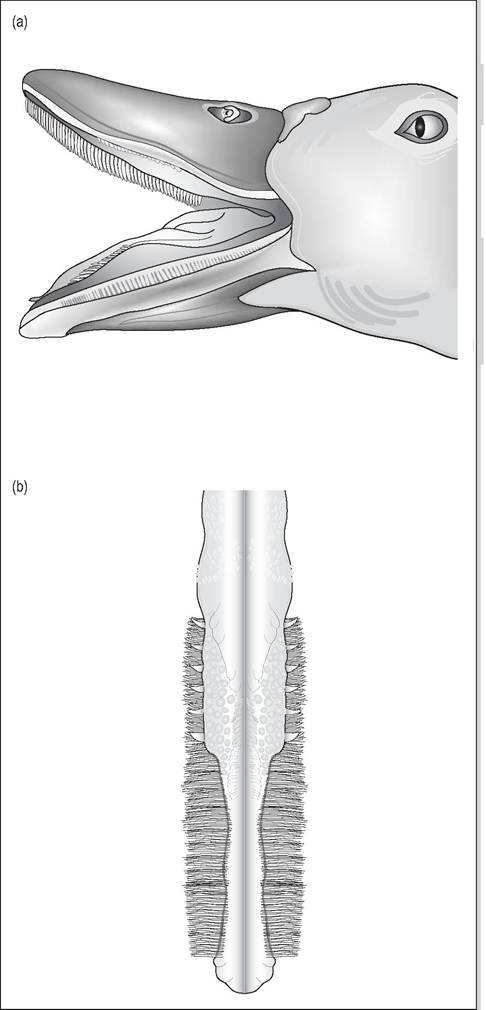
Figure 6.48 •
(a) Beak of duck showing upper and lower lamellae
(b) Tongue of duck showing lateral bristles - these bristles press against the lamellae to filter out the food particles
jection on the rostral beak called the egg tooth, which is used to pip the shell.
Cere
The base of the dorsal maxillary rhamphotheca is called the cere and it can be naked or fully feathered. It is composed of softer keratin than the rest and is particularly prominent and fleshy in waterfowl.
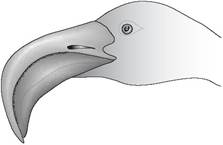
Figure 6.49 • The flamingo has a specialized bill that enables it to filter out its food from the water.
CLINICAL NOTE
The male budgie has a blue cere while the hen has a brownish pink cere. Male budgies however can develop a brownish cere under the feminizing influences of testicular Sertoli cell tumors.
Beaks are widely modified according to diet (Fig. 6.50). For example, finches have a stout conical bill for cracking seeds. In some species ridges on the hard palate serve to wedge the seed in place in order to crack open the shell with the lower beak. Hummingbirds have a long probing bill to penetrate deep into flowers for nectar. Raptors have hooked beaks for tearing prey while crows have a strong and pointed beak suited for their omnivorous diet.
Function of the beak
One of the main functions of the beak is to apprehend food and prepare it for delivery into the esophagus. However, the beak also has many other roles like preening, courtship, nest making, and the feeding of chicks.
In psittacines the beak is also used for locomotion and in these species the beak is connected to the skull by a synovial hingelike joint.CLINICAL NOTE
The powerful beak of psittacines is strong enough to crack a Brazil nut, so mind your fingers when examining large parrots.
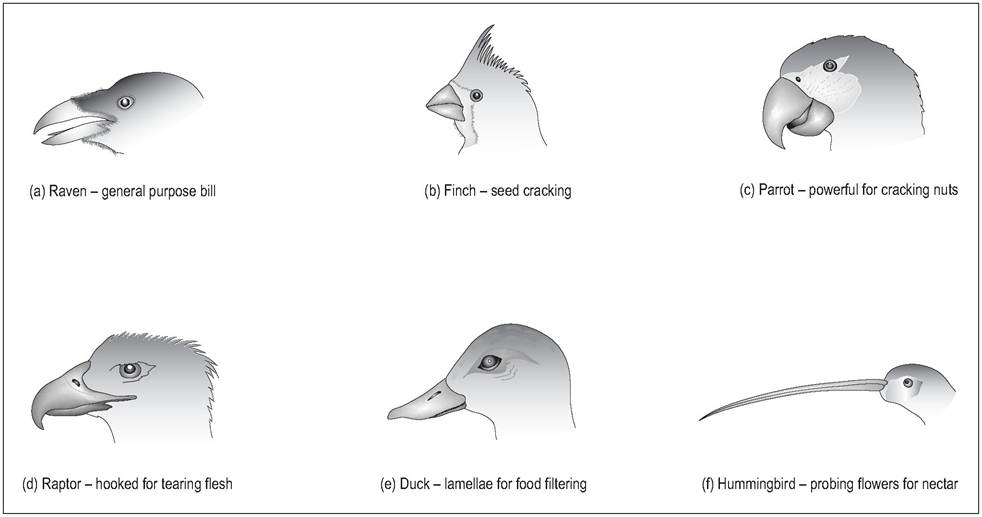
Figure 6.50 • Modifications of beak according to diet.
(a) Raven - general purpose bill
(b) Finch - seed cracking
(c) Parrot - powerful for cracking nuts
(d) Raptor - hooked for tearing flesh
(e) Duck - lamellae for food filtering
(f) Hummingbird - probing flowers for nectar
Oropharynx
In mammals the glossopalatine arches divide the oral cavity from the pharynx, and the soft palate divides the nasal cavity and oral pharynx. Birds, however, only have a common cavity called the oropharynx, which is often highly colored in newly hatched passerines to attract the parents attention while gaping for food (Klasing 1998). The hard palate is incomplete with a median slit called the choana, which links the nasal cavity to the oropharynx. This closes when food is swallowed to stop food entering the nasal cavity (Fig. 6.51). The oropharynx is lined by stratified squamous epithelium which is keratinized into papillae that line the oropharynx along the roof, choanal margins, infundibular cleft, and tongue to the laryngeal mound. These play a role in helping the food to pass caudally (King & McLelland 1984).
The infundibular cleft is a midline slitlike opening lying caudal to the choana, which is the common opening for the pharyngotympanic tubes (the avian equivalent of eustachian tubes). In birds these are short and not covered by folds in order to stop excess pressure building up in the middle ear when the bird flies (Hoefer et al. 1997; King & McLelland 1984). Lymphatic tissue called the pharyngeal tonsils lie in this cleft as well. The choana and infundibular cleft have a
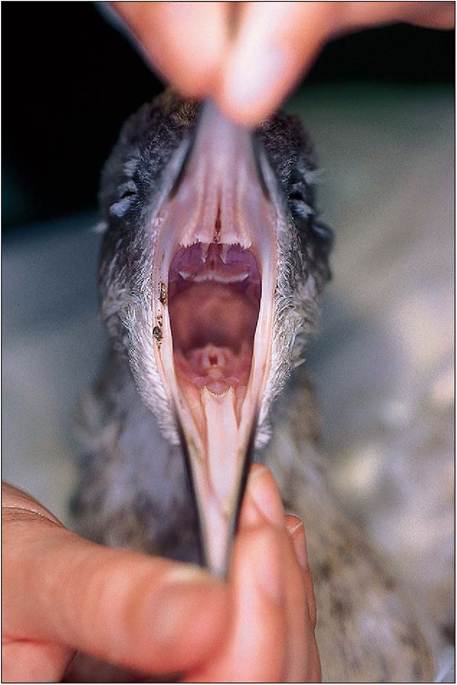
Figure 6.51 • Open mouth view showing choana, glottis and triangular tongue.
common opening in some birds like budgies (Melopsittacus undulatus) (Dyce et al. 2002).
CLINICAL NOTE
Many psittacines with Vitamin A deficiency develop blunted choanal papillae due to squamous metaplasia. These block salivary ducts and cause secondary bacterial infection; hence abscesses are commonly seen under the tongue and around the choana.
CLINICAL NOTE
When performing crop lavage under anesthetic it is important to block off the choana as well as inserting an endotracheal tube to avoid food entering the nasal cavity.
Tongue
The avian tongue is a highly diverse organ and can be used for both collecting, manipulating, and swallowing food, depending on the species (Table 6.4). In most species it is non-protrusible. It is supported by the hyobranchial (hyoid) apparatus, which consists of a rostral rod, the entoglossal bone, and a caudal rod, the rostral basibranchial bone. In most species the tongue itself is merely cornified mucous membrane covering the hyoid bone. Psittacines are unique in having paired entoglos- sal bones and a large muscular tongue capable of amazing dexterity (King & McLelland 1984).
Birds appear to have a poorly developed sense of taste, perhaps because of the rapid transit of food from mouth to crop. Taste buds are few in number and located on the palate, the base of the tongue, and the floor of oropharynx (see Senses).
Salivary glands
Birds have practically a layer of salivary glands scattered throughout the oropharynx. These are not well defined but there are small aggregates of maxillary and palatine glands in the roof of the mouth, and mandibular and lingual glands on the floor. Avian saliva is very different to the water secre-
| Table 6.4 Tongue types and functions (King & McLelland 1984) | ||
| Role of tongue | Type of species | Tongue type |
| Collecting food Manipulating food Swallowing food | Woodpeckers, hummingbirds Parrots, raptors, ducks, geese, swans Crows, domestic fowl | Protrusible Short, non-protrusible, caudal papillae Non-domestic fowl protrusible, caudal papillae |
GENERAL INTEREST
In some birds like woodpeckers the hyoid horns are so elongated they go completely around the back of the skull, ending near the upper maxilla.
This enables the woodpecker to protrude their tongue over four times the length of the upper beak to catch insects. Woodpeckers also have large mandibular glands beneath the tongue that secrete a sticky fluid to coat the tongue and act like flypaper to trap insects.tions of mammals as it is more mucoid to help lubricate food. Production of saliva is under the control of parasympathetic stimulation and is especially well developed in birds that eat a lot of dry food, such as insects or seeds (Duke 1986, 1993; Hoefer et al. 1997).
GENERAL INTEREST
130
Swifts secrete a mucilaginous substance from their mandibular glands, which is used like cement for their nests. In the Collocalia swift species of SE Asia the nest is built entirely of saliva and forms the gourmet Asian dish of bird's nest soup (King & McLelland 1984; McLelland 1979). tensible than in mammals. It is lined by stratified squamous epithelium with mucous glands to aid the passage of food down the tract. It contains only smooth muscles innervated by the vagus nerve, so excitement or fear can influence motility. Paralysis of the esophagus can occur in heavy metal poisoning causing stasis and impaction.
Regurgitation
This is when food is brought back by antiperistaltic movement of the esophagus. It is normal in species like pigeons and penguins while they are nursing neonates and is also used for sexual display in species like budgies.
Crop (Ingluvies)
This is a dilation of the esophagus located at the base of the neck just cranial to the thoracic inlet. It has the same epithelial structure as the esophagus but no mucus glands (Hill 1971a; King & McLelland 1984). In psittacines it is placed transversely and is easy to palpate as it is strongly attached to skin. The crop of pigeons is well developed and bilobed whereas waterfowl have a barely visible fusiform widening of the esophagus (Fig. 6.55).
CLINICAL NOTE
Feeding
The beak prehends the food, which is then lubricated by sticky mucus produced from the salivary glands. The lack of soft palate and pharyngeal muscles means that birds do not use peristalsis for swallowing. Instead papillae on the tongue and palate and rostrocaudal movement of the tongue moves the food into the caudal oropharynx. The head is then raised to let the food move passively, the glottis and choana close over and food gets flushed by the saliva into the proximal esophagus. From there it moves by peristaltic action down the gastrointestinal tract (Duke 1986, 1993; Hill 1971b; McLelland 1979).
Drinking
The beak is immersed in water and fluid enters the mouth by rostrocaudal movement of the tongue. Once in the caudal oropharynx the head is raised and it passes into the esophagus by gravity. Pigeons manage to suck water and do not have to raise their head while drinking; hummingbirds suck nectar through a tubular tongue. Psittacines can lap water due to their muscular tongue (King & McLelland 1984; McLelland 1979).
Esophagus
The esophagus lies to the right side of the neck, is thin walled and has longitudinal folds, which enables it to be more dis-
Crop impaction can be caused by ingesting large amounts of dry food. In raptors it can be blocked by the fur and feathers of prey if there is inadequate moisture in the diet (King & McLelland 1984).
The function of the crop in birds like psittacines and pigeons is to store food when the stomach is full (Evans 1996). In some birds, such as domestic fowl, harder food like grain is softened in the crop before traveling on to the proventriculus for physical digestion. In species like owls, gulls, and penguins, which have no crop, food is either stored in the esophagus or passes directly into the proventriculus. Food like fish can often be seen stretching from the proventriculus and sticking out of the beak in piscivorous birds without causing any choking or discomfort (Klasing 1998).
There is little chemical digestion in the esophagus and crop although salivary amylase may initiate carbohydrate digestion. The crop pH is acidic (4-6). It contains some gram-positive bacteria and a small amount of Candida species.
GENERAL INTEREST
The Hoatzin bird uses its extensive crop as a gizzard to help physically digest the leaves it feeds on. In these species the mass of the crop is 50 times that of the gizzard (Evans 1996; McLelland 1979).
Crop milk
Columbiformes produce crop milk to feed their young in the first few days after hatching. This is stimulated by prolactin and resembles milk, except it has no carbohydrate or calcium. It is produced in both sexes by desquamating cells from the stratified squamous epithelium of the crop and lasts until about 2 weeks after hatching. Psittacines, finches and Emperor penguins also produce crop secretions (McLelland 1979).
CLINICAL NOTE
The avian esophagus is very distensible so large tubes can be passed when gavage feeding to prevent the risk of accidental intubation of the trachea. For crop sampling, pass a tube into the crop and infuse some warm physiological saline. Aspirate a sample back and perform a wet smear and diagnostic cytology.
Stomach
In birds the stomach is divided into the glandular proventriculus and the muscular ventriculus (gizzard) divided by an isthmus. There are two types of stomach according to the diet of the species (Denbow 2000).
Carnivorous birds
These fish and meat eating species have little difference between the two stomachs, having a large easily distensible stomach to contain whole prey. The ventriculus in these species is used mainly for allowing time for gastric juices to act on the soft food rather than physical digestion (McLelland 1979).
Granivorous birds
These birds eat relatively indigestible foods so need a gizzard for grinding food. Consequently they have a thick-walled, non-distensible, and glandular proventriculus and a thick, muscular ventriculus (Figs. 6.56 and 6.57). Proventriculectomy is not easy in these species.
Proventriculus
This is the glandular portion of the stomach and its main function is production of the gastric juices and propulsion of food into the gizzard. It has two cell populations: epithelial cells that produce mucus and oxynticopeptic cells, which produce pepsinogen and hydrochloric acid (Denbow 2000; Hill 1971a). It lies to the left of the midline in the cran-
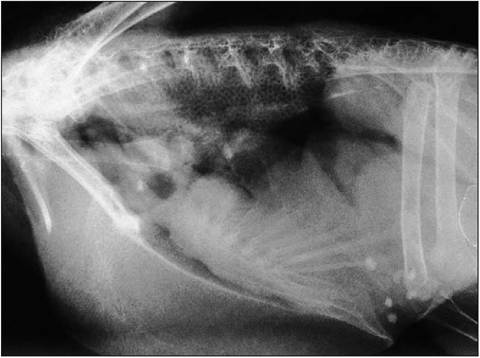
Figure 6.54 • Lateral radiograph showing normal honeycomb lungs, gizzard located at level of acetabuli and large pectoral muscle mass. This bird had renomegaly secondary to lymphoma.
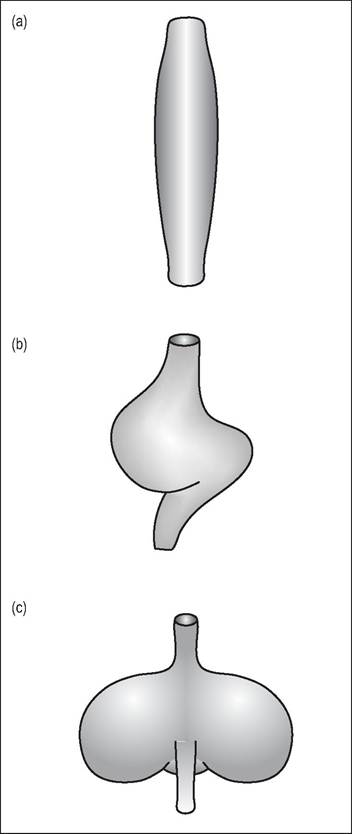
Figure 6.55 • The shape of the crop depends on the bird's diet.
(a) Waterfowl - simple and fusiform
(b) Parrots - seed can be softened here prior to passing into the ventriculus
(c) Pigeons - well developed and bilobed for softening grain and production of crop milk
iodorsal celom and is covered by other abdominal organs and the post-hepatic septum. There is no distinct esophageal sphincter and the loss of longitudinal folds is the only differing feature between esophagus and stomach.
Isthmus (Intermediate zone)
This is the transition from glandular stomach to muscular gizzard. It has no glands so the wall is thinner and less rigid than the proventriculus.
Ventriculus (Gizzard)
The ventriculus lies to the left of the midline and can be palpated as a firm mass just caudal to the sternum (Fig. 6.54). This second stomach is the site of protein digestion and
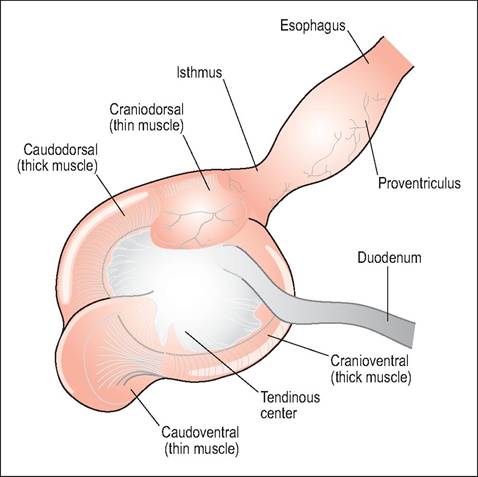
Figure 6.56 • External appearance of ventriculus (gizzard) in granivorous bird showing well-developed grinding muscles.
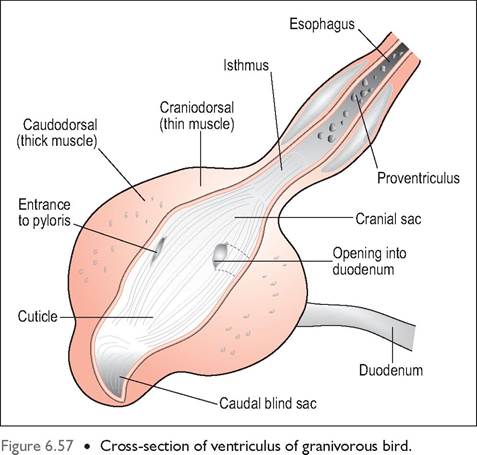
mechanical breakdown of food and consists of a large volume of smooth muscle lined internally by columnar epithelium. Koilin lines this epithelium like a cuticle and helps protect it during the grinding process. It is formed from mucosal cell secretions and is composed of protein and carbohydrate and becomes stained yellow-green due to bile reflux from the duodenum (Denbow 2000).
Herbivores (granivores) like turkeys and waterfowl tend to have a very developed ventriculus but it is less muscular in psittacine species that dehusk their seeds prior to swallowing (Klasing 1998; McLelland 1979).
CLINICAL NOTE
The ventriculus can be the site of foreign bodies so, if one has to be removed, the blind thin muscular sacs cranially and caudally are the best site for Ventriculectomy incisions. Waterfowl can also pick up lead weight which can be ground down by the contractions causing the lead to be absorbed.
The ventriculus has four muscular bands arranged so as to provide rotary and crushing movements to aid mechanical digestion. These consist of two thick layers of circular, smooth muscles that attach to a central tendinous aponeurosis on the left and right. They are dark colored due to the high myoglobin content of the smooth muscle. Cranially and caudally there are blind sacs formed by two layers of paler, thin muscles (King & McLelland 1984).
GENERAL INTEREST
The pulverizing action of the gizzard muscles is immensely powerful. In some bizarre medieval experiments performed on turkeys it was found that the gizzard could crush 24 walnuts in their shells in under 4 hours and turn 16 surgical lancets into grit in 16 hours!
CLINICAL NOTE
Psittacine and passerine species do not require grit for digestion as they crack open seeds and eat only the kernels. Chickens and pigeons swallow their seed whole and grind them in their gizzard using grit. Although grit is not essential in these species it markedly reduces gut transit times. Coarser food will require coarse grit (Duke 1986, 1993; Hoefer et al. 1997; McLelland 1979).
Food moves back and forth between proventriculus and ventriculus by contraction of the thick and thin muscles during stomach digestion. The opening of the duodenum lies very close to the proventriculus so that food not requiring grinding can bypass the gizzard completely.
Intestinal reflux
Intestinal reflux (also called egestion or casting) is performed by carnivores like Strigiformes (owls) and Falconiformes and is the method used to void indigestible portions from the ventriculus. It occurs due to synchronized contractions of the proventriculus, ventriculus, and duodenum about 12 hours post feeding. The presence of bulky food items in the stomach causes gastric contractions to increase and compact the pellet and push it into the distal esophagus. During reflux, gastric motility is inhibited and the pellet is expelled through the mouth by esophageal antiperistalsis.
The content and timing of the pellet depends on the bird's feeding habits. Owls eat their prey whole and have no crop for storage, so they egest bone as well as the usual roughage of feathers and fur after every meal. Falconiformes tear their food apart with their talons and, having crops, hold the food for longer and digest all the bone portion. They egest just once daily or at dawn as part of the circadian rhythm (McLelland 1979).
Pylorus
This lies to the right of the ventriculus. It is small in chickens but very large and dilated in cormorants and herons, perhaps to delay indigestible debris like fish bones for longer before entering the duodenum.
Liver
The liver is yellow color in a newborn chick because of the high fat content resulting from the absorption of yolk (McLelland 1979). This changes to the usual red-brown at around 10 days of age. It is proportionately large for the bird's size, compared to mammals. There are two main liver lobes, which surround the heart and join cranially at the midline. Some lobes are frequently subdivided. The liver is covered by a fat-filled post-hepatic septum. There is a well-developed hepatic portal circulation and the caudal vena cava enters the dorsal portion of the right lobe. The gall bladder lies on the visceral surface of the right lobe (King & McLelland 1984).
The gall bladder is absent in some Psittaciformes, Columbi- formes, and Struthioniformes (Bezuidenhout 1999; Evans 1996). In these species the bile flows from the right liver lobe to the ascending duodenum via the hepatoenteric duct.
Bile is important for avian digestion as it aids the emulsification of fats and also contains amylase and lipase. Many birds lack the enzyme bilirubin reductase so biliverdin is the main bile pigment (McLelland 1979). The liver synthesizes bile acids and empties them directly into the duodenum where they are reabsorbed and sent back to liver by entero- hepatic circulation. Therefore, hepatic malfunction can be measured by raised bile acids.
CLINICAL NOTE
A bird with liver problems will often pass urates stained green by the bile pigment biliverdin. Raised bile acid can also be a good indicator of hepatic disease.
CLINICAL NOTE
Some birds like Mynah birds and toucans store high levels of iron in their hepatocytes, leading to hemochromatosis or iron storage disease. The cause is unknown but it can be treated by recurrent phlebotomies to remove excess iron and by feeding a diet low in citrus fruits.
Pancreas
This is pale pink and lies within the antimesenteric border of the duodenal loop. It is composed of three lobes with one to three ducts that secrete the pancreatic enzymes into the ascending duodenum near the bile ducts. As in mammals, this gland has both endocrine and exocrine functions. It secretes the enzymes amylase, lipase, trypsin, and chymotrypsin and these help the breakdown of carbohydrates, fats, proteins, and bicarbonate. These are produced under the control of the hormone secretin and the vagus nerve (King & McLelland 1984).
Small intestine
In general, the small intestine is relatively shorter than in mammals but it is highly convoluted to help with birds' high metabolic demands. It is separated from the body wall by the caudal thoracic and abdominal airsacs. There is very little histological difference between the duodenum, ileum, and jejunum, which form a series of narrow u-shaped loops on the right side of the celom (McLelland 1979). The axial loop carries the Meckel diverticulum and marks the boundary between the ileum and jejunum, while the supraduodenal loop is the most distal part of the ileum (Evans 1996; King & McLelland 1984).
CLINICAL NOTE
The duodenum is easily seen on exploratory and necropsy as it lies most ventrally - caution must be taken to avoid incising it when entering the celomic cavity.
The bile and pancreatic ducts open into the distal part of the ascending duodenum opposite the cranial part of the gizzard. The jejunum and ileum are demarcated by the yolk sac remnant, the Meckel diverticulum. Altricial birds are fed by their parents so absorb the yolk sac speedily after hatching. Precocial birds have to fend for themselves from early on so rely on the yolk sac for nourishment during this period. It is normally absorbed from 10 days post hatching, leaving scar tissue behind.
The intestines are thin walled and narrow and consist of mucosa, submucosa, circular and longitudinal muscle, and serosa. The intestinal epithelium has folds and villi but, unlike in mammals, does not contain lacteals (Hill 1971a; McLelland 1979). Instead they absorb lipids through a well-developed capillary network. Three types of epithelial cell exist: chief cells, goblet cells, and endocrine cells. The chief cells have a brush border to absorb food, the goblet cells secrete mucus, and endocrine cells produce the hormones somatostatin, gastrin, and secretin. Although birds lack true mesenteric lymph nodes they have lymphoid nodules (Peyer patches) in the lamina propria to provide lymph drainage.
Large intestines
There is a short colorectum, which extends from the ileocecal junction to the cloaca. The colon is no thicker than the small intestine and reabsorbs water and electrolytes by antiperistaltic movements (Duke 1993). Urine is moved from the cloaca into the colon by this method.
Ceca
These are paired and arise at the junction of the ileum with the rectum. They run retrograde from the ileocecal junction with their blind end lying near the cloaca.
The mucosa has villi and there are scattered lymph follicles in the wall. The main function of the cecum is to aid the digestion of cellulose so they are most prominent in ground dwelling, flightless birds like ostriches and fowl (Bezuidenhout 1999; Hill 1971b). They are poorly developed in most arboreal birds, being rudimentary in pigeons and passerines and absent in raptors and psittacines (Duke 1993; Evans 1996; King & McLelland 1984). In fact more than two thirds of birds either have no ceca or have rudimentary ones (Klasing 1998).
Role of intestines and cecum
The small intestine is the principal site for chemical digestion. If ceca are present they contain gram-positive bacteria and protozoa which help to digest cellulose in herbivorous species.
Cecotrophy
Some species use cecotrophy to help survival on rough forage. Food passes down the intestines to the coprodeum by peristalsis. Occasionally an unknown mechanism returns the ingesta by retroperistalsis back up to the cecum. The long villi in the cecum separate the nutrient-rich food from the indigestible portion. High nutrient particles are absorbed and the rest is passed distally in a powerful cecal contraction to re enter the cloaca as cecal droppings. These semi-solid, chocolate-brown droppings are normally discharged about once a day (Duke 1986, 1993).
Cloaca
This is the site for termination of the urogenital and digestive systems. It is usually a bell-shaped dilation at the end of the rectum and consists of the copradeum, urodeum, and proctodeum (King, AS 1981a). The Bursa of Fabricius is located in the dorsal wall. The cloaca lies in the midline in mature males but in mature females the enlarged left oviduct pushes it to the right.
Coprodeum
This cranial compartment is where the rectum empties. This largest chamber is an extension of the colon. In some species it is lined by villi and is a site of water absorption. It is separated from the urodeum by the coprourodeal fold. If the rectum is full of feces this fold can bulge out of the vent dispelling feces without mixing them with urates. This fold can also close the coprodeum during egg laying to prevent feces being expelled at the same time (King & McLelland 1984).
Urodeum
This middle compartment is separated from the other parts by the circular mucosal folds. The ureters and genital ducts empty into its dorsal wall. The left oviduct opens into a small mound, which is covered by a small membrane in ducks, geese, and swans until the bird reaches sexual maturity. In male passerines the terminal vas deferens dilates during the breeding season to form a conical projection in the cloaca called the seminal glottus or cloacal promontory (Lake 1981).
Proctodeum
This is a short compartment separated from the urodeum by the uroproctodeal fold. It empties contents into the vent, which is controlled by the external anal sphincter. The Bursa of Fabricius is located in its dorsal wall. It is the site of B-lymphocyte production and differentiation, which are necessary for humoral immunity. Male ducks, geese, and swans have a large proctodeum to house the large pro- trusible phallus on the ventral floor.
Vent
This is the external opening of the cloaca. In psittacines this is a circular opening, but it is transverse in domestic fowl and ‘u'-shaped in ducks and geese. The sphincter muscle has an outer and inner circle of striated muscle. There is also a transverse muscle that originates from the pelvic bone, passes ventral to the proctodeum, and helps pull the vent into position for copulation.
KEY POINTS
Modifications of the avian gastrointestinal tract:
• Teeth replaced by horny beak
• No soft palate so single large oropharynx
• Open pharyngotympanic (eustachian) tubes
• Esophagus is on the right side of the neck and length and diameter are greater than in mammals
• Lots of mucoid saliva produced to lubricate food
• Crop stores and softens food
• Two stomachs - proventriculus for chemical digestion and ventriculus for grinding
• Oxynticopeptic cells in stomach produce hydrochloric acid and pepsinogen
• Columbiformes and Galliformes require grit to help grind down hard food
• Biliverdin is the main bile pigment of birds - hence green urates with liver problems
• Cloaca is the common termination of digestive and urogenital tracts