RESPIRATORY SYSTEM
Birds use their respiratory system not only for gaseous exchange but also for vocalization and thermoregulation.
The avian respiratory system is unique in that the lungs are small, undergo little change in volume when breathing, and have airsacs that act as bellows but do not participate in gas exchange.
This segregation of ventilation and gas exchange helps to increase the total gas exchange surface area. The bellows system allows continuous gas flow as opposed to the “in and out” tidal flow of mammals. The net result is that in birds up to 10 times more oxygen can be absorbed.Birds have no diaphragm, instead the horizontal septum separates the lungs from the viscera. This septum plays no active role in respiration but passively helps to displace the viscera during breathing.
Nasal cavity
Birds can breathe through their nose or mouth (Powell 2000). The nasal cavity is compressed laterally and divided medially by a very thin septum. The nasal openings or “nares” lie laterally at the base of the beak, where they can be surrounded in species like psittacines by a thick cere. Many species also have a hard, keratinized structure called the operculum at the center of each nare that acts as a baffle to prevent inhalation of foreign bodies. Air enters through these nares into the conchae of the nasal cavity, passes down through the slitlike choana and across the oropharynx to the glottis (King & McLelland 1984).
CLINICAL NOTE
Some birds, like gannets and boobies, have no external nares but breathe through a narrow gap between the distal upper and lower beak. This enables them to dive at high speed without water blowing out of their nostrils (Fig. 6.36).

Figure 6.36 • Juvenile gannet (Sula bassana) showing lack of external nares.
Nasal conchae
The conchae are highly vascular, epithelial folds in the nasal cavity that increase the surface area over which air flows, allowing it to be filtered and warmed. They are divided into rostral, middle, and caudal parts and play a major role in olfaction, filtering, and thermoregulation (King & McLelland 1984).
Olfactory epithelium in the caudal conchae picks up scent particles, ciliated epithelium filters out foreign particles, and mucus secreted by goblet cells flushes these through the choana into the oropharynx. An extensive network of blood vessels, the rete mirabile, helps control the water and heat loss by warming or cooling air entering the nasal cavity. This helps prevent excessive water loss and is particularly important in desert and migrating birds.
Infraorbital sinus
This paranasal sinus is particularly well developed in psittacines and becomes superficial ventromedial to the orbit (Fig.
CLINICAL NOTE
Nebulization is an excellent way of rehydrating an avian patient and helps compensate for any evaporative water loss due to respiratory problems.
6.37). It has numerous diverticula, which extend into the premaxilla, around the ear and rostral orbit, and into the lower beak. It communicates dorsally with the caudal and middle conchae and also with the cervicocephalic airsac at its caudal extent. In some birds (Psittaciformes and Anseriformes) the right and left sinuses communicate.
Nasal gland
The nasal or salt gland lies just dorsal to the orbit and opens into the nasal cavity at the level of the rostral concha from where saline is then sneezed from the nostrils. It secretes a hypertonic solution enabling seabirds to drink seawater. The gland is similar to the kidney and has a countercurrent blood supply which removes sodium ions from the bloodstream (King & McLelland 1984).
Larynx
The laryngeal opening (or rima glottis) is slitlike and, unlike in mammmals, is not covered by an epiglottis (Fig. 6.51).
It is usually found at the back of the tongue but is often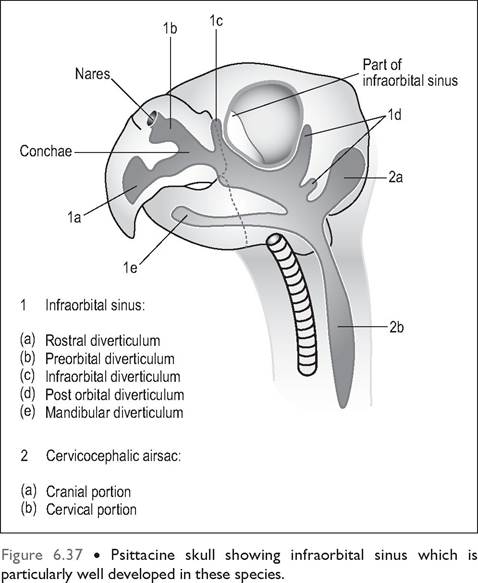
CLINICAL NOTE
In psittacines the free communication between the nasal cavity, infraorbital cavity, skull, and airsacs makes the treatment and drainage of sinus infections very difficult. Often the first signs of infection can be swellings rostral and ventral to the eye where the infraorbital sinus lies superficially just under the skin and subcutaneous tissues (Fig. 6.38).
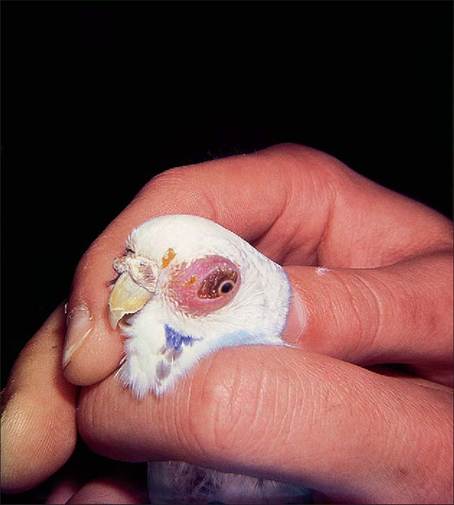
Figure 6.38 • Swelling of the infraorbital sinus in psittacines is a symptom of sinusitis and not an ocular problem.
To compensate for the increased tracheal length, birds minimize air resistance by having a trachea of wider diameter than mammals of equivalent size. Birds with tracheal convolutions and greater lengths also have wider diameters to increase total volume. However, this results in birds having a dead space 4.5 times that of mammals (Hinds & Calder 1971). To overcome this birds have increased tidal volume and (relative to size) a slower and deeper rate of breathing (Hinds & Calder 1971). For example, a mammal of 100 g would have a resting respiration rate of 100 breaths per minute while in a bird of equivalent size it would be 30 per minute (Welty 1982b). Birds with long necks, like swans, breathe as slowly as 10 breaths per minute and the ostrich has the slowest rate of all birds at 3 to 5 breaths per minute (Brackenbury 1987).
Syrinx
The syrinx (Greek word for panpipe) is the bird's voice box and is analogous to the mammalian larynx except that it lacks vocal cords which vibrate with air currents. It is rudimentary in species like vultures and ostriches (King, AS 1989).
The syrinx can be classified as tracheal, tracheobronchial or bronchial depending on location. The most common type is the tracheobronchial (Fig.
6.39), located at the bifurcation of the trachea, and has a median cartilage called the pessulus, at the bifurcation. This type is found in psittacines although they lack a median pessulus.The syrinx consists of a series of modified tracheobronchial cartilages, two vibrating tympaniform membranes and muscles which vary the membrane tension. These thin membranes line the medial and lateral bronchi and sound is produced during expiration by vibration of air through the
obscured by the fleshy tongue in psittacines. There is no soft palate but the rima glottis regulates the passage of air by a dilator and constrictor muscle that prevents aspiration of food material. The larynx plays no role in sound production (King & McLelland 1984).
Trachea
The trachea is composed of rigid interlocking rings and is relatively long, given birds' size. This is because birds need long necks to help manipulate objects with their beaks. The greater air resistance created by elongation means the diameter of the rima glottis and trachea is wider to compensate (Hinds & Calder 1971).
In some species the trachea is so long it forms coils; in the Trumpeter swan these lie in a cavity of the sternum. Toucans and mynahs have a normal ventral deviation of the trachea just cranial to the thoracic inlet. In penguins, a median septum divides it into left and right tubes, making it very easy to intubate a primary bronchus by mistake (King & McLelland 1984; Powell 2000).
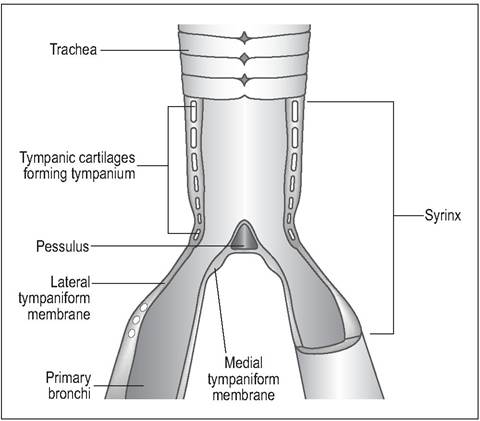
Figure 6.39 • Longitudinal section of tracheobronchial syrinx showing tympaniform membranes and syringeal cartilages.
syrinx (King, AS 1989; King & Molony 1971). The surrounding interclavicular airsac gives the voice resonance by pushing against these membranes. Birds with long coiled tracheas have deep resonant voices, hence the call of the Trumpeter swan (Gaunt 1987).
The number of syringeal muscles varies with species.
Despite their great speaking ability parrots have a relatively simple syringeal apparatus with only three pairs of syringeal muscles (Evans 1996; Welty 1982b). Hawks have only one pair while the songbirds have five pairs (King, AS 1989). In many songbirds the two sides of the syrinx are independently controlled allowing them, literally, to sing a duet with themselves (Gaunt 1987; King & McLelland 1984).CLINICAL NOTE
As the trachea is narrowed at the site of the syrinx this is a common site of obstruction by foreign bodies like seeds or fungal granulomas. Birds exhibiting voice changes should have the syrinx examined endoscopically. Other common causes of voice changes can be goiter pressing on the syrinx or damage to the surrounding interclavicular airsac.
CLINICAL NOTE
Male ducks have modified their tracheal syrinx into an osseous bulla to give their voice resonance. This can be seen radiographically near the left side of the thoracic inlet on the ventrodorsal view and can be used in sexual differentiation in monomorphic species (Dyce et al. 2002; King, AS 1989).
Lungs
The avian lungs are far more rigid than mammalian lungs because they contain more cartilage and are thus less compressible. They are small, compact and spongy and, being quite dense, occupy less space than mammalian lungs. They are bright pink in color and closely adherent to the rib cage dorsally, which leaves furrow marks on the surface. Ventrally they are lined by the horizontal septum. Due to the absence of a diaphragm the lungs do not collapse if the celom is entered via surgery or endoscopy (Fig. 6.40).
Bronchi
Each primary bronchus runs through the whole length of the lungs (where they are called mesobronchi) and terminates in the caudal airsacs. On entering the lungs each bronchus then gives rise to four sets of secondary bronchi named according to the area of the lung they supply: mediodorsal, medioventral, laterodorsal, and lateroventral (Fig.
6.44). These terminate in tertiary bronchi called parabronchi where blood gas exchange takes place. All bronchi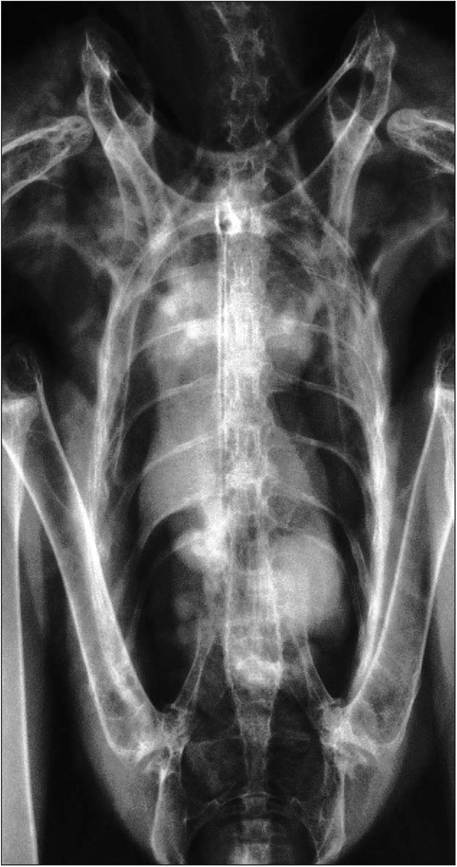
Figure 6.41 • Ventrodorsal radiograph of lungs and airsacs in a normal bird of prey.
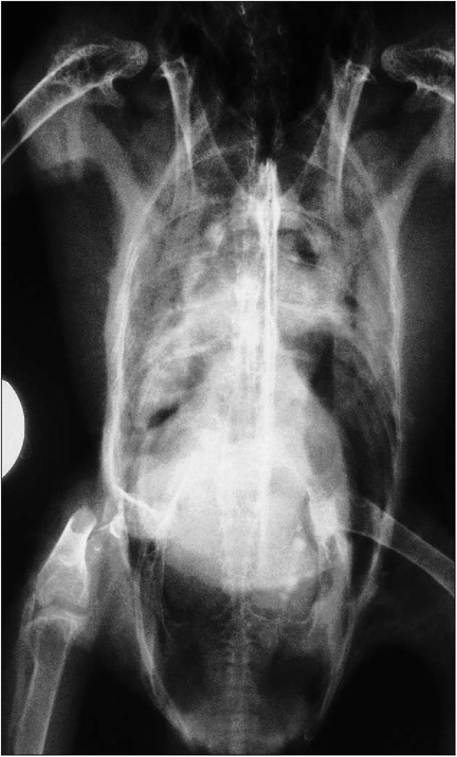
Figure 6.42 • Ventrodorsal radiographs of lungs and airsacs in a parrot attacked by a dog, showing fractured femur and hemorrhage in the thoracic airsacs.
CLINICAL NOTE
During anesthetics tidal volume may be reduced by as much as half when the bird is lying on its back. This is due to compression of the viscera on the caudal air sacs (King & Payne 1964) (Fig. 6.43). The absence of a diaphragm also means that, when anesthetized, birds may not breathe regularly, owing to the lack of sternal muscle contraction. Ventilation by positive pressure ventilation is always recommended (Fedde 1993).
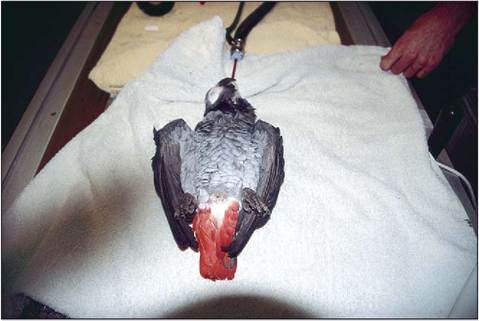
Figure 6.43 • Prolonged dorsal recumbency should be avoided during general anesthetics due to mass of the viscera compressing caudal airsacs.
CLINICAL NOTE
On lateral radiographs the tertiary parabronchi are seen end on and this gives a honeycomb appearance to the lung (Fig. 6.54). Absence of this normal appearance can indicate pathology (Smith & Smith 1997).
are lined by smooth muscle causing dilation and contraction (Lasiewski 1972).
Parabronchi
The parabronchi, which can be seen by the naked eye, make up the bulk of the lung tissue. They have invaginations called atria which lead to a labyrinth of microscopic air capillaries extending out to perform gas exchange (Fig. 6.45). These lie in close proximity to tiny blood capillaries, giving the lung a highly vascular appearance. The air capillaries are lined by surfactant, which keeps small cavities open by lowering surface tension. If this is not present, surface tension at the air-blood interface will collapse the capillaries leading to build up of transudate fluid (Lasiewski 1972; Powell 2000).
The number of these tertiary parabronchi increases with active flight; for example ducks have approximately 1800 whereas the flightless domestic fowl (Gallus gallus) has only 300 to 500 (Scheid & Piiper 1987).
Most of these parabronchi are a parallel series of hundreds of tubes called paleopulmonic bronchi. In most birds* there are also irregular branched parabronchi (“neopulmonic bronchi”), which never comprise more than 25% of the parabronchi (Maina 1996; Powell 2000; Scheid & Piiper 1987). Air flow through the paleopulmonic is always in the same direction during inspiration and expiration; however, in the neo- pulmonic bronchi it is bi-directional according to the phase of breathing. As no airway valves have been found it is thought that the shape and alignment of the secondary bronchi creates a pressure differential between the cranial and caudal airsacs influencing air flow through the parabronchi (Brackenbury 1987; McLelland & Molony 1983; Scheid & Piiper 1971).
*Some more primitive birds, like the kiwi, penguin, and emu have only paleopulmonic bronchi and these do have bi-directional flow (Fedde 1993; King & McLelland 1984).
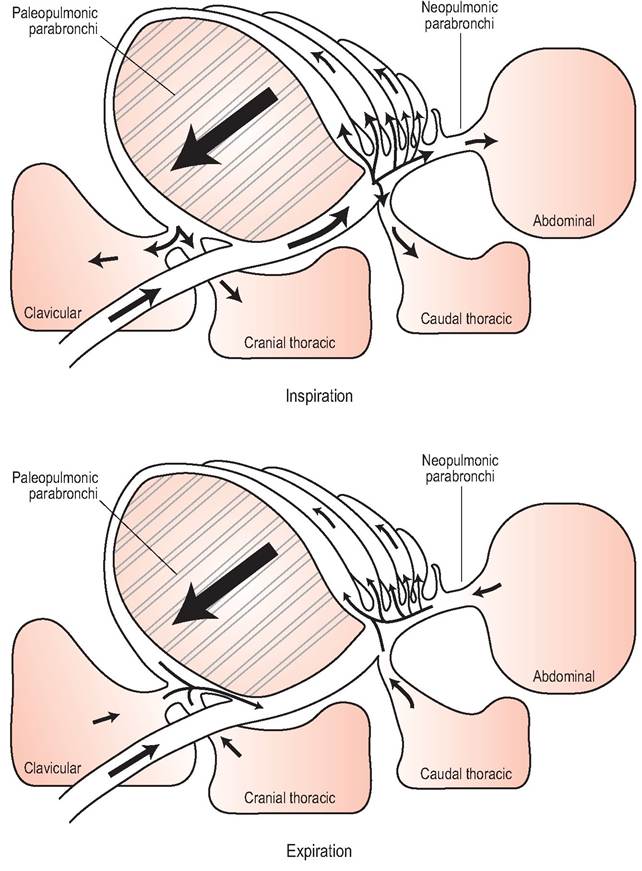
Figure 6.44 • Diagram showing air flow through the respiratory tract of birds during inspiration and expiration. Air flows in the same direction from caudal to cranial in both phases of respiration.
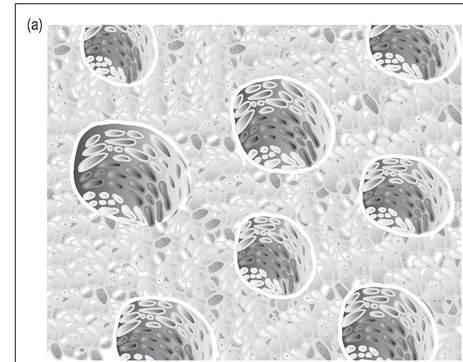
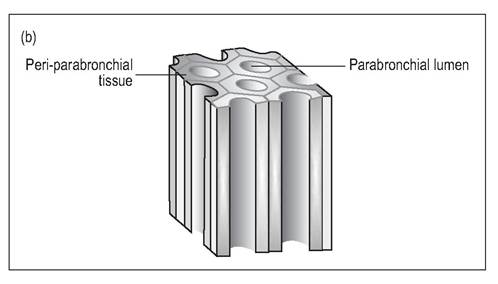
Figure 6.45 •
(a) Cross-section of the tubular parabronchi showing air capillaries where gas exchange between blood and inhaled air takes place.
(b) Longitudinal section of parabronchi.
Gaseous exchange
Flight and the ability to fly at altitude means that birds have much higher oxygen demands than mammals. Avian lungs are 10 times more efficient than mammalian lungs in capturing oxygen due to the following modifications:
Thin blood-gas barrier
The air capillaries of the lung (equivalent to the mammalian alveoli) are finer and more numerous and the blood-gas barrier is very thin (Evans 1996; King & Molony 1971; Scheid & Piiper 1972). In fact, the capillaries are so narrow it is said the erythrocytes have to line up in single file! This is possible because, unlike mammalian lungs which have to expand and contract with every breath, the fixed avian lungs require little interstitial tissue for added strength. Stronger fliers like the passerines have a thinner blood-gas barrier than flightless birds such as chickens (Maina 1989; McLelland & Molony 1983).
Cross-current blood flow
The blood flow is at right angles to the airflow, giving a cross-current exchange system. This means that blood flow is always at right angles no matter which way the air is flowing (Scheid & Piiper 1972, 1987). Cross-current exchange allows more efficient absorption of oxygen without incurring high levels of carbon dioxide in the blood (Fedde 1993; Maina 1989; McLelland & Molony 1983).
One way airflow
The airflow through the lungs is unidirectional as opposed to the “in and out” tidal flow of mammals (Scheid & Piiper 1972) (Fig. 6.46). The parabronchi, being tubes and not deadend sacs like alveoli, allow for continuous gas exchange in the avian lung and this may explain why birds can fly at high altitudes (Maina 1989; Scheid & Piiper 1987; Schmidt- Nielsen 1975, 1990).
Rigid lung
The fact that the lungs are rigid and play no role in ventilation means that there is 20% more area for gas exchange than
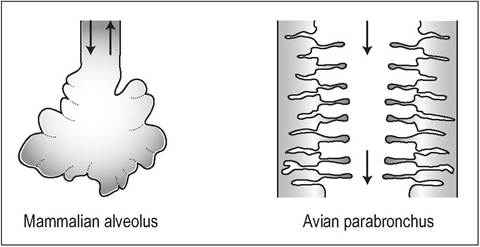
Figure 6.46 • Comparison between mammalian “to and fro” ventilatory system via alveoli and the “flow through” avian system of parabronchi.
in mammals. However, this rapid efficiency of gas exchange can have its downside as it makes bird much more susceptible to inhaled toxins and infections.
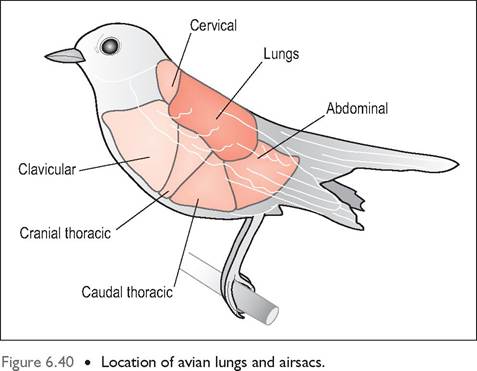
Airsacs
These are thin (two cells thick), distensible and transparent sacs, lined by simple squamous epithelium. They constitute 80% of the respiratory volume and extend from the body cavity into the wing, vertebrae, and leg bones (Lasiewski 1972; Maina 1996). There is little blood supply as they play no role in gas exchange (Dyce et al. 2002; Fedde 1993). In most birds there are three pairs of airsacs and two single ones, creating eight in total (Figs. 6.41 and 6.42). In some species the cervical sacs are paired, making nine airsacs (Scheid & Piiper 1987). Each airsac is connected to the secondary bronchi at an area called an ostium which is usually located along the ventrolateral border of the lung. The parabronchi also interconnect with all the airsacs except the cervical one.
The airsacs are usually divided into two groups according to their connections to the bronchi: the cranial airsacs (cervical, clavicular, and cranial thoracic), which connect to the ventral bronchi, and the caudal airsacs (caudal thoracic and abdominal), which connect to the primary bronchus (Fig. 6.44).
Function of airsacs
The airsacs function to create a unidirectional flow of air through the lungs, which is important for maximizing oxygen extraction. They help to reduce the amount of heat produced during flight by evaporation from the airsacs. They also play a role in sound production, courtship displays in some species, and possibly cooling of the testes for spermatogenesis.
Cervical airsac
This lies between the lungs, dorsal to the esophagus and has vertebral diverticula from C3 - T5. It can be paired in some birds. It also communicates with the skull via the infraorbital sinus. It may help to provide buoyancy and insulation and reduce the point of impact in diving birds.
Clavicular airsac
This lies in the thoracic inlet and has two parts: the intrathoracic diverticulum around the heart and sternum, and the extrathoracic, which spreads between the bones of the pectoral girdle and into the proximal humerus. This sac extends into the syrinx and is essential for sound production.
CLINICAL NOTE
Compound fractures of the humerus can introduce infection via the pneumatic bones into the lungs and airsacs.
Cranial and caudal thoracic airsacs
Both these sacs are paired and lie in sequence in the dorsolateral thoracic cage. The caudal thoracic airsacs are the only ones which lack a connection with pneumatic bones.
Abdominal airsac
This is paired and reaches from the lungs to the cloaca. It has diverticula around the kidneys and femora. It is potentially the largest in all birds. In the live bird this sac can be entered during laparoscopy, close to the ostium at the angle between the lateral border of the ilium and the caudal border of the dorsal end of the last vertebral rib.
HOW DO BIRDS BREATHE?
Birds do not have a diaphragm so the air sacs act like a bellows to move air through the non-expansile lungs. This is controlled by the action of the muscles, which act independently during flight.
Inspiration
There are six muscles of inspiration but the principal ones are the external intercostals and the costosternalis. On inspiration the rib cage moves outwards and the sternum moves downward. This negative pressure causes air to be sucked in via the nares to the air sacs. Half of this will enter the caudal airsac via the neopulmonic bronchi and half will enter the cranial airsacs via the paleopulmonic bronchi. No air passes directly into the cranial airsacs (McLelland & Molony 1983).
Expiration
There are nine muscles of expiration, the main ones being the internal intercostal and the abdominal musculature. On expiration the rib cage moves upwards, reducing chest size and compressing air from the airsacs back through the lungs. Air from the caudal airsacs passes through the paleopul- monic bronchi into the cranial airsacs. Air from the cranial airsacs flows out of the primary bronchi and trachea (McLelland & Molony 1983).
Respiration
Respiration rate
The smaller the bird the faster the respiration rate. It can vary from 40-50 breaths per minute in cockatiels to 25-30 in pigeons and 13 in turkeys. The ostrich with 3-5 breaths per minute has the slowest rate of all (Brackenbury 1987). Flying birds increase their respiration rate by 10-12 times above their resting rate, rising to 20-30 times with maximum exertion.
Muscles of respiration
Respiration is controlled by the external and internal intercostal and abdominal muscles. Birds have the same abdominal muscles as in mammals although, as the sternum is so large, they are smaller. Unlike mammalian ventilation, which takes one respiratory cycle, it takes two ventilation cycles to move air right through the avian system. This does not mean the two cycles are different but that it would take a single bolus of air two breaths before it can be exhaled (McLelland & Molony 1983).
Both inspiration and expiration require active muscle contractions and airflow in the paleopulmonic parabronchi is always in a caudal to cranial direction.
CLINICAL NOTE
The avian flow-through system means that artificial ventilation can be achieved by fresh oxygen down the trachea or via an airsac cannula if the trachea is obstructed. Manual compression by compressing and lifting the sternum can also be undertaken (Powell 2000).
KEY POINTS
• Birds have no diaphragm.
• The trachea is longer and much wider than in mammals, creating more dead space.
• The location of the syrinx at the bifurcation of the trachea means this is commonly a site for foreign body blockage.
• Birds have increased their oxygen absorption by separating gaseous exchange from ventilation, having a thin blood-air barrier, cross-current exchange, and a flow-through system.
• Airsacs function solely in ventilation; as they are poorly vascularized the caudal airsacs easily harbor infections.
• Airsac cannulation can be life saving in cases of obstructive dyspnea.
CLINICAL NOTE