» External Features and Integument
Feathers provide the principal characteristic that distinguishes birds from mammals. They streamline the body and assist in transforming the forelimbs into wings. The feathers are among the features (others are mentioned later) that lighten birds relative to their size and thus enhance their efficiency in the air.
Feathers have many functions that in mammals are performed by hairy skin: thermoregulation, communication, and protection against mechanical, radiological, thermal, chemical, and biological influences.The skin is thin and loose and tears easily. Because of poor vascular and nerve supply, the skin wounds do not bleed as much as in mammals, and birds seem relatively insensitive to manipulation of their skin. The skin in chickens is yellowish over the body but may be more deeply pigmented on the shanks and feet. It is paler in productive laying hens, in which the pigment is withdrawn and incorporated in the yolk. The dorsal surface of the neck-trunk junction, the cranial skinfold of the knee, and the lateral side of the thorax are recommended for subcutaneous injections. In most species, including the domestic chicken, localized changes in the skin occur during the brooding period for the more efficient incubation of the eggs. Brooding (incubation) patches that develop on the breast are characterized by feather loss and by thickening, edema, and increased vascularity. The subcutaneous layer is mainly composed of loose connective tissue. It also contains fat, most copiously present in aquatic and arctic species such as penguins, ducks, geese, and swans, and in migratory species before migration.
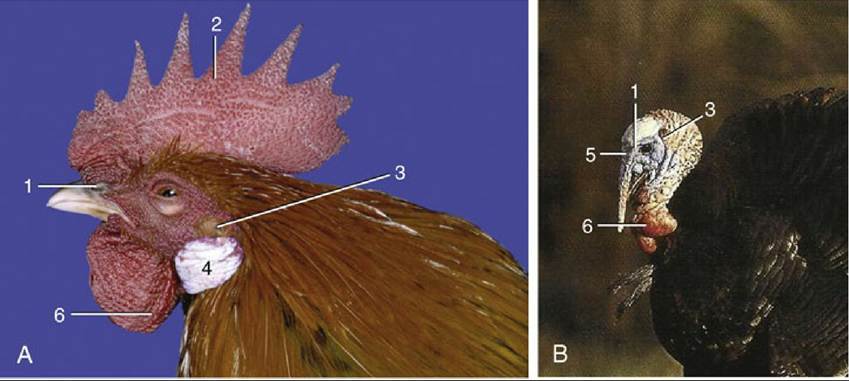
FIG. 37.1 Head of the chicken (A) and the turkey (B). 1, Nostril; 2, comb; 3, ear opening; 4, earlobes; 5, snood; 6, wattles.

FIG.
37.2 (A) and (B), Two chickens with ornaments. 1, Comb; 2, wattles; 3, earlobe.The comb, wattles, and earlobes (and the snood of turkeys) are soft ornamental outgrowths of the skin about the head (Figs. 37.1 and 37.2). Their dermis is thick and vascular, but the covering epidermis is thin. They are thus easily injured and provide potential portals for infection. In nearly all commercially reared chicks, the comb (and snood) are snipped off (dubbing, desnooding) to prevent their traumatization in the confined spaces in which these birds are held. The edges of the wattles are used for intradermal injections.
The beak (bill) is the functional skin-derived counterpart of the lips and teeth of mammals. It provides a horny cover (rhamphotheca) for the rostral parts of both upper (rhinotheca) and lower (gnathotheca) jawa and grows continuously to compensate for natural wear. The beak varies tremendously in form among species, according to diet (Fig. 37.3). A rich innervation causes it to be quite sensitive. Most commercially raised chickens and turkeys are debeaked when young (cutting off the upper beak in front of the nostrils) to prevent cannibalism. In psittacines, pigeons, and raptors, the base of the maxillary rhamphotheca, called the cere, may enclose the nostrils (Fig. 37.3C and D).
The beak is composed of softer keratin than the rest and is particularly prominent and fleshy in waterfowl as well as in budgerigars, in which it is used as a guide to their sex. The cere of the cock is blue, and that of the hen is light brownish pink.

FIG. 37.3 (A to D) Differences in the form of the avian head. (E) The filter mechanism in the beak of a duck.
The scales on the shanks and feet are cornified epidermal patches similar to those of reptiles (Fig.
37.4). The feet of most birds are adapted for perching or holding prey and have one toe facing backward and three facing forward (anisodactyl).
In waterfowl the three forward-pointing toes are webbed to make more efficient sculls (palmate). Some species, such as psittacines, have two (first and fourth) toes facing backward and two (second and third) facing forward (zygodactyl) to enable grasping and climbing. The spur with an osseous core within a cone of horn, on the caudomedial surface of the rooster's shank is used as a weapon. The length of the spur and the growth rings at its base may be used for determining age. Removal of the spur papilla in the chick inhibits its growth, much as the removal of the horn bud prevents horn growth in ruminants.There are only three discrete skin glands: the sebaceous uropygial gland (preen or oil gland; Fig.
37.5), the aural gland, and the vent gland. The absence of sweat glands means that the birds have to lose heat through their skin and by evaporation from the respiratory system. The epidermis has a unique feature that allows it to act like a holocrine sebaceous gland, secreting a thin lipid film that helps in the maintenances of the plumage.
The uropygial gland in chickens is bilobed, about 2 cm in diameter, and located dorsal to the vertebrae that form the short tail. Its fatty secretion that emerges from paired openings atop a small cutaneous papilla is carried to the body and wing feathers during preening. In waterfowl the secretion is important for waterproofing the feathers and insulating the submerged part of the body. This bacteriostatic lipid layer may be the reason why birds are little prone to skin infections. The uropygial gland is prominent in budgerigars and African grey parrots (Psittacus erithacus) but absent from many other parrots (e.g., Amazon parrots), ostriches, and many pigeons.

FIG. 37.4 Left foot of a cockerel (A) and a goose (B). 1, Shank (metatarsus); 2, spur; 3, web between toes; I to IV, toes.
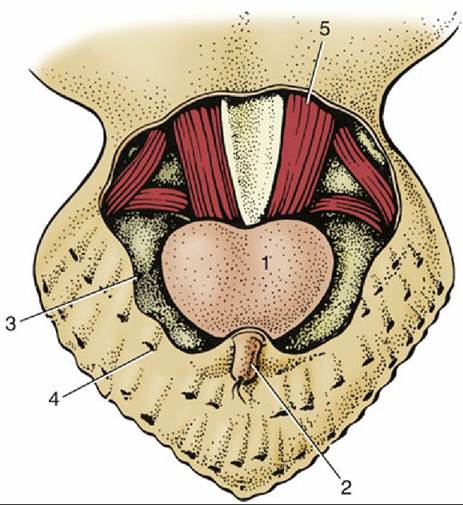
FIG.
37.5 Uropygial (preen) gland, dorsal view (schematic). 1, Uropygial gland; 2, papilla of uropygial gland through which the secretion is extruded; 3, cut edge of skin; 4, feather follicle; 5, caudal vertebrae and associated muscles.Aural sebaceous glands around the external ear secrete a waxy substance. Vent glands secrete mucus; their function is uncertain but may be linked to internal fertilization.
The Feathers
Feathers are highly specialized epidermal structures that have evolved from the scales of reptiles. Although light in relation to their size, they are of sturdy construction. Six types are recognized (contour feathers, semiplumes, filoplumes, down feathers, afterfeathers, and bristles), but only the contour and down feathers are described here. The contour are the externally visible feathers of flight that modify the body contours, the wings, and the tail. The contour feathers (tectrices) conceal the down feathers, which create an effective dead air space that insulates the body. The wing feathers (remiges) are made of approximately 10 primaries or hand feathers (Fig. 37.6) and 10 to 20 secondaries or arm feathers. The tail feathers (rectrices) are attached to the pygostyle and are used for steering and braking during flight. There are usually 6 pairs, but numbers can vary from 4 to 10 pairs depending on the species. Feathers are concentrated in tracts (pterylae), leaving bare areas (apteria) that are preferred surgical sites. Feathers hide emaciation.
The exposed portion of a typical contour feather consists of a main shaft extended on each side by the vane (Fig. 37.7). The vanes in wing feathers are asymmetrical; the external side of the vane is narrower than the internal for aerodynamic flight. The vane consists of numerous closely ranked branches (barbs; Fig. 37.7/2) that leave the shaft at angles of about 45 degrees. Adjacent barbs are connected by large numbers of minute barbules to form the level surfaces of the vane. This connection is effected by microscopic hooks on the distal ranks of barbules that loosely engage the proximal barbules crossing under them (Fig.
37.7/3'). Neighboring barbs are easily disconnected but reattach if brought together, as in preening or grooming the feathers.
FIG. 37.6 Wing feathers of a pigeon. There are 10 primary or hand feathers and 10 secondary or arm feathers.
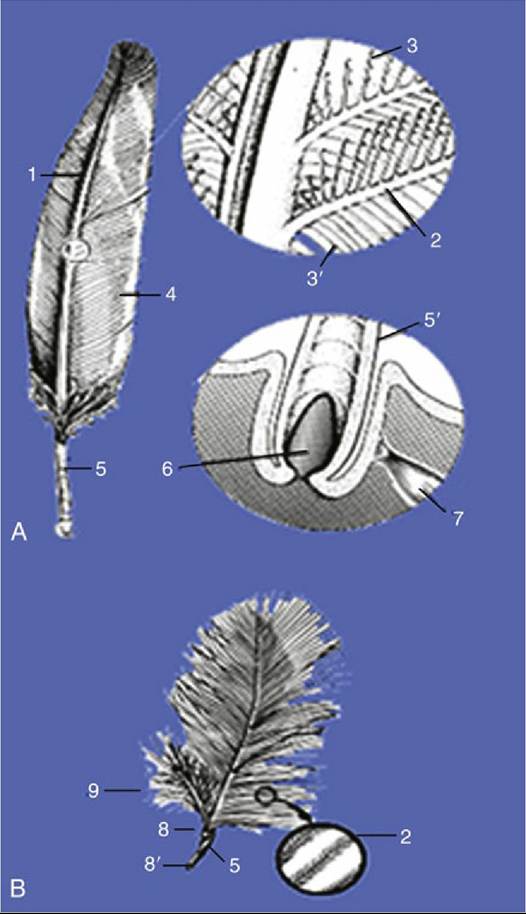
FIG. 37.7 (A) Contour feather (with enlargements). (B) Down feather (with enlargement). 1, Main shaft; 2, barb with barbules; 3, distal barbules with microscopic hooks; 3', proximal barbules; 4, vane formed by the barbs; 5, quill; 5', quill in feather follicle; 6, dermal papilla; 7, feather muscle; 8, distal umbilicus; 8', proximal umbilicus; 9, afterfeather.
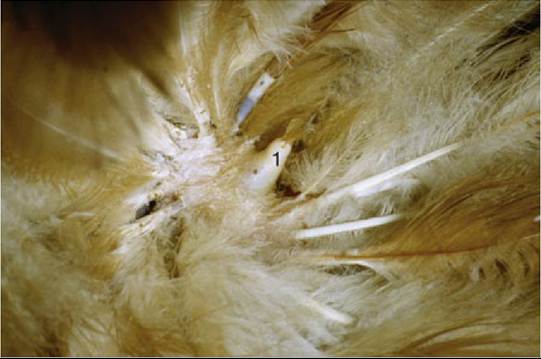
FIG. 37.8 Dermal papilla (1).
The main shaft on the undersurface of the feather presents a longitudinal groove that ends in a depression (distal umbilicus; Fig. 37.7B/8) opposite the fluffy proximal part of the vane. A small downy afterfeather (hyopenna; Fig. 37.7/9) may emerge from the umbilicus and contribute to the fluffiness.
The embedded part (quill, calamus) of the feather occupies the feather follicle, an oblique tubular invagination of the skin (Fig. 37.7/5'). The small dermal papilla at the bottom of the follicle extends into the opening (proximal umbilicus) at the proximal end of the quill (Fig. 37.8). The quill itself is hollow and contains air and cellular debris (pulp caps) derived from the papilla. Feather muscles (Fig. 37.7/7), similar to the mammalian arrector pili muscles, attach to the sides of the follicles; they often form extensive networks that elevate or lower whole groups of feathers.
The barbs of the down feathers (Fig. 37.8) do not interlock to form a closed vane. Their haphazard arrangement gives these feather their fluffy appearance. In pigeons and many psittacines, such as cockatoos and African grey parrots, specialized down feathers produce a fine powder keratin dust from the barbs.
This talclike powder is then coated over the plumage during preening; its absence is often the first sign of psittacine beak and feather disease. In pigeons the production of powder down (bloom) has been associated with human allergic alveolitis or pigeon fancier's lung.Feather color plays a major role in camouflage, courtship, and protection from heat and light. Color, produced by pigments and by the intersection of light with feather structure, may complement other features of the feathers or feather tracts in indicating sex. Other species are monomorphic, and in them sex determination is dependent on endoscopy or molecular techniques (DNA analysis).
Colors of Feathers: The black tyrosine-derived pigment melanin, which also produces grays and browns, is the most commonly found in birds. The red, orange, and yellow carotenoid pigments that produce such colors as cardinal red and flamingo pink are obtained from the diet. Blue pigments are not found in birds, but the color may appear when white light is scattered by feathers that absorb the red (short-wavelength) end of the spectrum while the blue end is reflected (Tyndall effect). More green is produced by the combination of this effect with a yellow carotenoid pigment than is produced by porphyrins. Porphyrins, nitrogenous pigments also synthesized by birds, provide green, red, and some browns. They occur in gallinaceous birds, pigeons, and owls and may fluoresce when exposed to ultraviolet light. The iridescence seen in starlings and peacocks is produced by a combination of melanin with the structural breakdown of light striking the feather barbules. The color varies with the angle from which it is viewed.
At set times birds replace their feathers (molt or ecdysis) to discard worn ones or to change their plumage for display or camouflage. This occurs usually once a year after the breeding season (postnuptial or winter plumage) and is regulated through the actions of the thyroid hormone. Other factors influencing molting are nutrition, time of year, temperature, and light. It also depends on habitat and whether the species is migratory. Young birds molt their juvenile feathers before they become adults and often go through a series of subadult plumages. During molt, which is a slow and gradual process, birds should be provided rest and a diet rich in protein (especially the amino acids lysine, cystine, and arginine) and minerals (calcium and iron) to support the higher metabolic demands (increases of 15%-25%) made by the rapid epidermal proliferation and loss of insulation. Birds in poor condition often produce misshapen feathers. In most species, replacement of the large contour (flight) feathers is sequential (inside primaries first) and symmetrical so that flight always remains possible. Ducks and geese, however, lose all of these feathers at once, leaving them temporarily flightless. The old feather is pushed out by epidermal growth at the base of the follicle, and as it vacates the follicle, its replacement begins to grow. Before the barbs are released, they are encased in a sheath called a bloodfeather or pinfeather. The loss of a feather by plucking initiates a similar sequence of events. Clipping feathers is therefore unlikely to permanently disable birds for flight.