General anatomy
The optic nerve is actually a tract of the CNS as it is myelinated by oligodendroglia, thus the term ‘nerve’ is a misnomer (pl). It is the only part of the CNS that can be observed on a neurological examination.
Photoreceptors (rods and cones) are the receptors of the visual system and convert light into receptor potentials. Rods are highly sensitive to light, therefore they are used during dim light conditions, while cones are less sensitive to light but respond to specific light frequencies, that is, colours. The presence of 20 times more rods than cones in the canine retina accounts for dogs’ good night vision. The vertebrate retina is inverted in the sense that the light-sensing cells sit at the back of the retina, so that light has to pass through layers of neurons before it reaches the photoreceptors. This arrangement ensures that incoming light only stimulates photoreceptors once. If the photoreceptors were at the front of the retina, then some of the incident light would stimulate them, while the remaining light could pass to the back of the retina. There it would be reflected back into the eye at a new angle stimulating different photoreceptors. Thus light from a single visual point would stimulate photoreceptors in different regions (incident versus reflected) causing loss of resolution and visual blurring.
There is a three-stage neuronal system in the retina. The photoreceptors form the deepest layer, the middle layer contains integrating neurons, while the superficial layer contains the multipolar ganglion neurons, the axons of which pass across the retina to collect at the optic disc and form the optic nerve. Thus light passes through the cornea, anterior chamber, iris, posterior chamber, ganglion and bipolar layers to reach the photoreceptors. Receptor potentials from the rods and cones undergo complex processing by integrating neurons of the retina but ultimately result in action potentials in retinal ganglion cells.
Several important features of visual perception can be traced to the retinal encoding and processing of light.In most domestic mammals, axons of retinal neurons are unmyelinated until they coalesce at the optic disc. Thus the opaque nature of myelin does not interrupt the passage of light through the retinal layers to the photoreceptors. However, there are no photoreceptors at the optic disc, resulting in a blind spot in the visual field. Myelination of the optic nerve fibres makes the optic disc appear creamy white on fundic examination. In some species, such as rabbits, there is some myelination of retinal ganglion axons. This may be observed clinically as a white streak extending from the optic disc.
The optic chiasm and decussation
The optic nerve extends caudally from the retina, through the optic foramen in the presphenoid bone into the neurocranium. It joins the ventral aspect of the diencephalon at the optic chiasm, just rostral to the hypophysis (pituitary gland). In general, the majority of axons decussate, but the degree of decussation depends on the type of animal. In fishes and birds, all fibres decussate (Fig. 10.10). In mammals, there is partial decussation (ungulates about 80-90%, dogs 75%, cats 65%, primates 50%).
Light also crosses in the lens of the eye, thus axons decussating at the optic chiasm come from the nasal retina and temporal field of view, while the temporal retinal axons receiving the nasal field of view, remain ipsilateral (see Fig. 10.5).
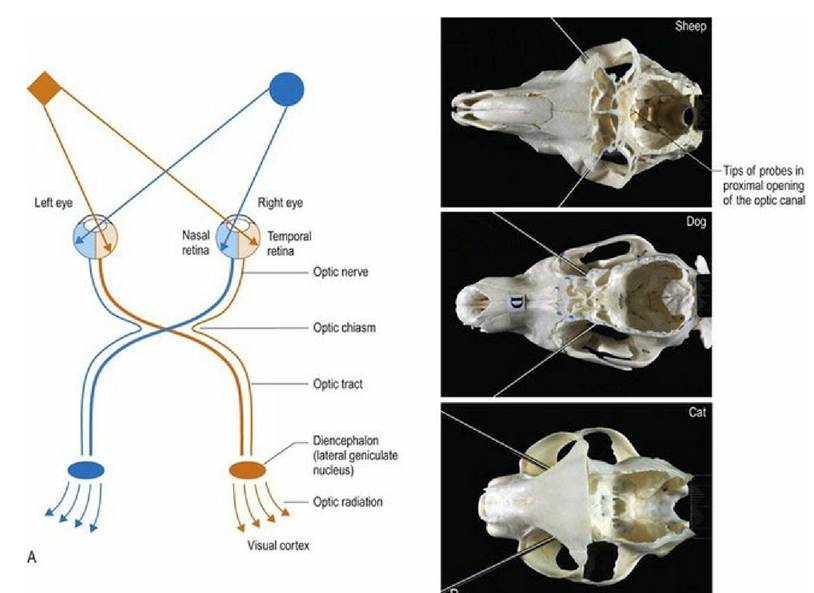
Fig. 10.5 (A) The optic pathway and transmission of light stimuli to the visual cortex. (B) Examples of
optic axes of the sheep, dog and cat, identified using metal probes placed in the optic canals. The angles between the axes for the sheep, dog and cat are 100°, 70° and 58°, respectively; the decreasing angles indicate increasing overlap of the visual fields.
The visual pathway continues caudal to the optic chiasm as the optic tracts.
In animals in which there is partial decussation at the optic chiasm, each optic tract contains axons from the contralateral nasal field and the ipsilateral temporal field. Thus the optic tract contains axons from retinal areas of the two eyes that are receiving input from the same part of the visual field (Fig. 10.5A).The placement of the eyes and decussation of the optic chiasm is functionally significant. Animals with laterally placed eyes, e.g. ungulates/prey animals (Fig. 10.5B) have wide fields of view and a high percentage of fibres decussate at the optic chiasm. Thus the majority of visual input is processed in the contralateral visual cortex. A wide field of view permits these animals to see behind them; this is useful to detect danger such as stalking predators. But depth of field/binocular vision requires processing of visual input from both eyes in the same visual cortex, thus prey species have poor binocular vision. Conversely, animals with eyes that face forward, like the cat, have smaller fields of view, but have better binocular vision due to decreased decussation at the optic chiasm. This results in the animal receiving input from each eye on both sides of the brain. For example in Fig. 10.5A, input from the orange diamond located in the left side of the visual field will project to the nasal retina of the left eye and the temporal retina of the right eye. The input from those retinal areas will both end up in the same visual cortex - that is the right side one. Thus the same region of the brain processes two sets of information seen by the separate eyes, about the same object. This dual perspective results in binocular vision which is essential for depth perception. Depth perception is required for accurate judgement of distance and how fast an object is moving, therefore it is essential for predatory animals. The ability to perform coordinated, conjugate eye movements, including convergence (focussing both eyes on the same object) is also greater in animals with forward-facing eyes and reduced decussation.
The total field of view combined from both eyes in the horse is approximately 320°. That comprises a field of view for each eye being about 200°s, with about 65° of overlap. The overlapping visual field is especially well developed ventrally for viewing objects on the ground. However, there is not overlap immediately in front of the nose, creating a blind spot. Most dogs have a total visual field of 250°. The degree of binocular overlap is about 75° for long-nosed dogs to 85° for short-nosed breeds. Comparatively, humans have a maximum total horizontal field of view of approximately 180°, 120° of which comprises the binocular field of view, flanked by two uniocular fields of approximately 40°.
Post-chiasmatic visual pathways
From the optic chiasm, axons course Caudodorsolaterally in the optic tracts. The majority of axons connect to lateral geniculate nucleus (Fig. A17) of the caudal diencephalon (metathalamus) and then via the optic radiation to the visual cortex for visual perception. Alternatively, axons travel to the midbrain for reflex function (see Visual reflexes). Using the cat as an example, approximately 80% of fibres from each eye go to the lateral geniculate nuclei and 20% to the midbrain.
The visual cortex is divided into specific functional regions. In primates, different areas process input regarding stationary objects, spatial relationships and depth perception, or panoramic vision for movement. The visual cortex also makes connections to adjacent association cortex, motor cortex, midbrain and cerebellum. Connections to the midbrain are essential for normal vision.
Lesions in the visual system rostral to the chiasm (pre-chiasmatic lesions) can result in blindness in one eye, since the optic nerve consists of fibres from only one eye. Post-chiasmatic lesions however, can result in changes in vision from both eyes as the optic tract contains fibres from both eyes.