Hepatic Metabolism Switches from Glucose Utilization to Glucose Production During the Postabsorptive Phase
As the absorption of a meal is completed, the rate of glucose absorption from the gut wanes, and the blood glu∞se concentration diminishes, removing the stimulus for insulin production.
As blood glucose concentrations decline, glucagon secretion is stimulated. The primary target organ of glucagon is the liver, in which glucagon creates marked metabolic changes. Through stimulation of specific cell surface receptors on hepatocytes, glucagon activates adenyl cyclase, leading to the phosphorylation of numerous cellular enzymes (see Chapter 1). Some enzymes are activated by phosphorylation, whereas others are inactivated, and unless the overall scheme of substrate flow is considered, the whole phosphorylation-dephosphorylation system appears to be quite random and to make little sense. Considering the actions of the individual enzymes in light of their effect on the flow of energy substrate through the liver, however, reveals that the system is an elegant and incredibly well- orchestrated mechanism for the maintenance of fuel homeostasis.The enzymes that stimulate mobilization and utilization of fuels are activated by phosphorylation, whereas those stimulating storage of fuels are inactivated by phosphorylation. It must be understood that many enzymes of intermediary metabolism serve a passive role, catalyzing reactions that can go in either direction, depending on substrate concentrations. A relatively small number of regulatory enzymes usually stand at the head of metabolic pathways and determine the substrate concentrations to which the other, unregulated enzymes are exposed. Through its effect on several key regulatory enzymes, glucagon (a stimulator of phosphorylation) places the liver in a fuel-mobilization state. In contrast, insulin (an inhibitor of phosphorylation) promotes a hepatic metabolic pattern that favors fuel storage, as previously discussed.
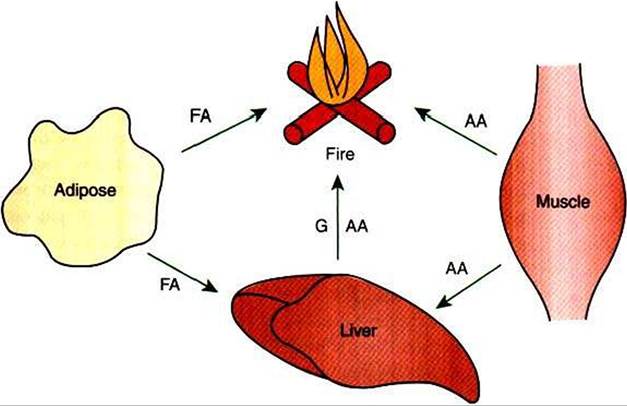
FIGURE 32-11 Postabsorptive metabolism is characterized by movement of fuels out of depot sites for immediate use. Glucose (G) arising from glycogenolysis or gluconeogenesis is a major fuel, although some fatty acid (FA) is consumed as well. Amino acid (AA) forms the substrate for gluconeogenesis.
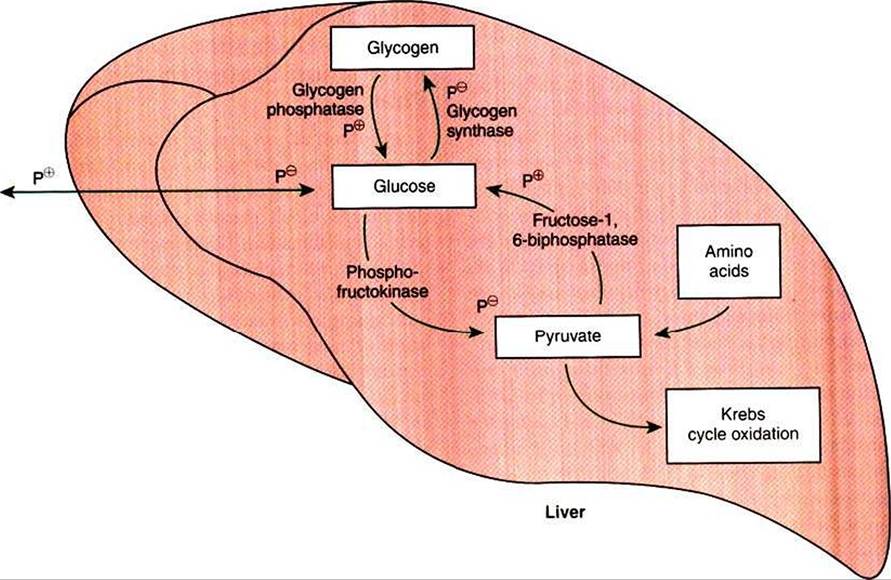
FIGURE 32-12 Effects of phosphorylation on four key enzymes of glucose production and utilization. All four enzymes are phosphorylated under the influence of cyclic adenosine monophosphate (cAMP). Note, however, that the enzymes that favor glucose formation are stimulated by phosphorylation (P*), whereas those that favor glucose utilization and storage are inhibited by phosphorylation (P^).
The opposing actions of insulin and glucagon on hepatic metabolism are evident from their actions on two key regulatory enzyme pairs: glycogen synthase and glycogen phosphatase, «and phosphofructokinase and fructose- l,6-bisρhosρhatase. The first of these pairs regulates glycogen synthesis and breakdown, whereas the second regulates glycolysis and gluconeogenesis, respectively. Figure 32-12 illustrates the actions of these enzymes and their regulatory effects. GIycogen synthase and phosphofructokinase are inhibited by phosphorylation and thus are stimulated by insulin. Glycogen phosphatase and Iructose- 1,6-bisphosphatase are stimulated by phosphorylation and thus stimulated by glucagon. The actions of insulin and glucagon on these antagonistic enzyme pairs emphasize the importance of the insulin∕g)ucagon ratio to which the liver is exposed. Neither hormone elicits an “all-or-none” reaction, but rather alters the balance of opposing reactions by influencing the relative activity of antagonistic enzymes. Thus the fuel-mobilizing or fuel-storing activity of the liver depends on which hormone is most dominant.
For this reason, the insulin/glucagon ratio appears to be more important to liver metabolism than the absolute concentration of either hormone.Under the influence of glucagon, glycogen phosphatase is activated by phosphorylation, promoting glycogenolysis and the elevation of intracellular glucose concentrations. As glucose accumulates, it is prevented from cycling back into glycogen because the major enzyme catalyzing that reaction, glycogen synthase, is blocked by phosphorylation. In addition, the flow of glucose into glycolysis is also blocked by phosphorylation inhibition of phosphofrιιctokinase (Figure 32-12). Thus the normal pathways for glucose utilization within the hepatocyte are all inhibited by glucagon, allowing glucose from glycogen breakdown to accumulate in the cells. Eventually, intracellular glucose escapes into the extracellular fluid and on into the blood. In this manner, hepatic glycogen is mobilized to elevate and maintain blood glucose concentrations when they begin to decline.
The liver stores of glycogen are relatively limited and cannot maintain blood glucose concentrations for a long period. Estimates in humans are that hepatic glycogen will serve blood glucose needs for 6 to 12 hours under conditions of light exertion and for only about 20 minutes under conditions of heavy exertion. Values for animals are probably similar. Therefore, in addition to glycogen mobilization, some other means must exist for maintaining the body’s glucose supply during periods of exertion and prolonged periods between meals. Under these conditions of greater demand, glucose is provided by gluconeogenesis. Gluconeogenesis is promoted by the phosphorylation-stimulated enzyme fructose- 1,6-bisphosphatasc. This enzyme essentially puts the glycolytic pathway into reverse, leading to glucose production from the same molecules that are intermediates in its oxidative destruction. Important substrates are pyruvate and all the intermediates of the Krebs cycle.
At this point, it is important to remember that most of the Krebs cycle intermediates or pyruvate can be supplied by the deamination of amino acids. The entry point of the various amino acids into the scheme of carbohydrate metabolism is illustrated in Figure 32-8. Pyruvate and all the Krebs cycle intermediates can flow backward through the oxidative pathway (not all the reactions of gluconeogenesis are the exact reverse of the corresponding reactions in glycolysis, but the net result of gluconeogenesis is the reverse of glycolysis), resulting in the production of glucose. Thus, amino acids provide a large store of precursors for glucose formation. The end result of glucagon stimulation is to promote the production of glucose through glycogenolysis and gluconeogenesis, turning the liver into a glucose-synthesizing organ.