Hormones of the Adenohypophysis
Growth Hormone
The release of growth hormone (GH), also called somatotropin or somatotropic hormone, from the adenohypophysis is regulated by hypothalamic factors that either stimulate (GH- releasing hormone, or GHRH) or inhibit (GH release-inhibiting hormone, GHIH, or somatostatin) release.
GH levels are highest in young, growing animals, but adult animals continue to secrete it. increases in GH secretion in adults occur in response to a variety of stimuli, but probably the most important physiologic stimulus is a reduction in plasma glucose. in adults, GH functions as a regulator of metabolism during starvation, deficits in plasma glucose, or hibernation. it acts to reduce protein breakdown and the use of glucose for energy in skeletal muscle and to increase the mobilization of fatty acids from adipose tissue.The role of GH in the determination of body stature in growing animals was introduced in Chapter 5. As was discussed earlier, GH itself has little direct effect on cartilage proliferation and bone growth in young animals. its growthpromoting effects are mediated by other peptides, somatomedins (primarily insulin-like growth factors 1 and 2 [IGF-1 and IGF-2]), which are released by the liver and cells in the area of growth plates in bone when stimulated by GH. The somatomedins are the direct stimulators of chondrocytes within the growth plates. Somatomedins also have negative feedback effects on the hypothalamus and adenohypophysis to regulate the release of GH.
in addition to GH, the secretion of somatomedins by the liver of young growing animals is regulated in part by nutrition. inadequate nutrition may retard growth in part because of a suppression of somatomedin secretion. small dogs have lower blood levels of IGF-1 than large dogs, suggesting that within a species body size and IGF-1 levels are correlated.
Excessive GH in young animals leads to gigantism. Increases in body size are not possible in older animals where growth plates are closed. Excessive GH (with associated IGFs) in mature animals leads to acromegaly. Further increases in stature are not possible in mature animals, but cartilage proliferation around joints and other skeletal locations produces enlargements in these areas and a characteristic coarseness of facial features. Affected adults also have derangements of carbohydrate and lipid metabolism (increased blood levels of glucose and fatty acids) because of the metabolic effects of excessive GH.
The amino acid sequence of GH varies among mammalian species, and thus GH produced by one species is not always biologically effective in a different species. Recombinant DNA technology has been used to produce both human recombinant GH and recombinant bovine somatotropin. The human product is used clinically to prevent certain types of human pituitary dwarfism, and the bovine product is used to increase both lactation and food efficiency.
Adrenocorticotropic Hormone
The primary target cells of adrenocorticotropic hormone (ACTH) are the cells of the adrenal cortex (outer region of the adrenal glands) that produce glucocorticoids. Glucocorticoids are steroid hormones that function in the regulation of metabolism (discussed later).
Adrenal Glands. The two adrenal glands are located close to the kidneys (ad, toward; ren, kidney). shape, size, and exact location vary from one species to another. Each gland consists of an outer region, the adrenal cortex, and an inner region, the adrenal medulla. These parts of the adrenal gland come from separate embryonic precursors and have distinctly different functions, in spite of their close physical association within a single connective tissue capsule.
The blood supply to the adrenal gland varies, but in general, small arteries enter the capsule surrounding the gland. These arteries are derived directly from the aorta or from its branches, including the renal, intercostal, and lumbar arteries.
Veins from the adrenal gland drain to the caudal vena cava.Three zones or regions of the adrenal cortex can be identified by light microscopy in most mammals, and each zone is the source of different hormones. From outermost to innermost, the three layers or zones are zona glomerulosa, zona fasciculata, and zona reticularis (Fig. 12-5). All hormones secreted from all three zones are steroid hormones, so cells of
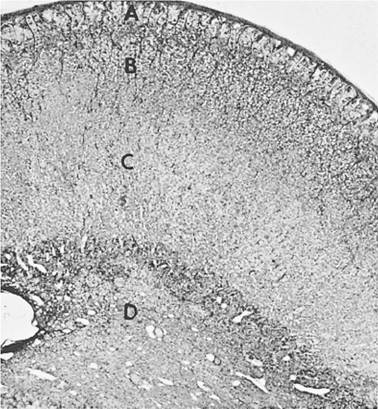
Figure 12-5. Adrenal gland of the horse. The cortical zones (A, zona glomerulosa; B, zona fasciculate; and C, zona reticularis) can be recognized, as can the medullary region, D. (Reprinted with permission of Wiley- Blackwell from Dellmann H.D. Veterinary Histology: An Outline Text-Atlas. Philadelphia: Lea & Febiger, 1971.)
all zones have ultrastructural characteristics of steroid-secreting cells.
The hormones secreted by the adrenal medulla (epinephrine and norepinephrine) are amines and are stored in secretory granules prior to release. These endocrine cells are termed chromaffin cells because of their affinity for chromium stains. As was described in Chapter 10 in the section on the autonomic nervous system, epinephrine and norepinephrine are released from the adrenal medulla in times of stress, and this release is regulated via the autonomic nervous system. A tumor of chromaffin cells is termed a pheochromocytoma, and this neoplasia typically results in excessive secretion of epinephrine and norepinephrine. The resulting clinical signs are those of excessive stimulation of the sympathetic nervous system (e.g., increased heart rate and blood pressure and increased metabolic rate).
Hormones of the Adrenal Cortex. The zona glomerulosa secretes mineralocorticoids (primarily aldosterone) that function in the regulation of sodium and potassium balance. The regulation of balance is primarily accomplished by controlling the loss of sodium and potassium in urine; more details on their specific functions are given in Chapters 18 and 23.
Mineralocorticoid secretion is not regulated by ACTH but rather by the serum potassium concentration and the renin-angiotensin system, another group of chemical messengers discussed in more detail in Chapter 18.Glucocorticoids (primarily cortisol and corticosterone) are the major secretory product of both the zona fasciculata and zona reticularis, and ACTH is the major regulator (stimulator) of their secretion. Without ACTH the fasciculata and reticularis both atrophy, but the glomerulosa remains intact. The inner zones of the adrenal cortex are also a source of adrenal sex hormones (androgens and estrogens), but the rates of secretion in normal adult animals are very low and not necessary for normal reproductive behavior and function.
The secretion of ACTH from the adenohypophysis is stimulated by a hypothalamic hormone, corticotropin-releasing hormone (CRH), and this is the most important regulator of ACTH release. However, the regulation of CRH, hence ACTH release, is extremely complex and affected by a wide variety of stimuli. ACTH increases are considered to be a classic sign of stress, and plasma levels of ACTH or cortisol are often used in experimental settings to evaluate the overall stress placed on an animal by any type of physical or emotional stimulus (e.g., restraint, starvation, presence of a predator). Both ACTH and glucocorticoids have negative feedback effects on the pituitary and the hypothalamus to maintain normal resting blood levels of ACTH and glucocorticoids, but stressful stimuli can override these effects.
Glucocorticoids have many target tissues throughout the body. in general their effects on these target tissues would seem an appropriate response to counteract stressful stimuli. For example, glucocorticoids increase the rate of gluconeogenesis (glucose formation) by the liver and increase the rate of fatty acid mobilization from lipid tissue. in skeletal muscle protein synthesis is reduced and protein degradation is increased, which means that more amino acids are available for gluconeogenesis by the liver. These metabolic effects are particularly important during starvation.
Glucocorticoids are often used therapeutically to inhibit inflammatory and immune responses. The doses used for these effects produce blood levels that are much higher than those seen in normal animals, even when they are responding to stress. Such levels and effects are described as supraphysiologic or pharmacologic. Among the many components of the inflammatory process that are inhibited by glucocorticoids are the synthesis pathways for prostaglandins, leukotrienes, and thromboxanes.
Thyroid-Stimulating Hormone
The target cells for thyroid-stimulating hormone (TSH), also called thyrotropin, are endocrine cells of the thyroid that produce and release thyroxine
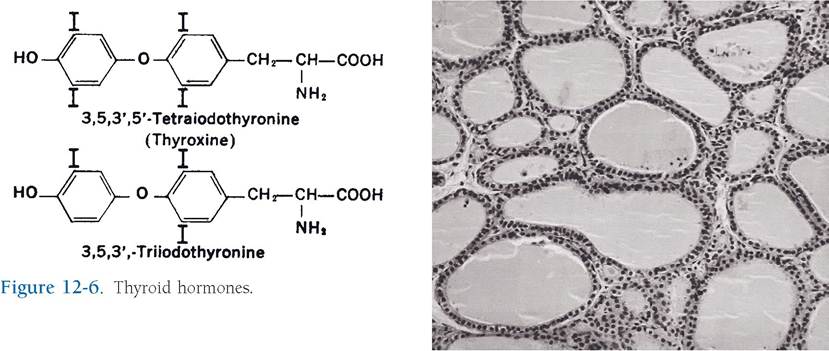
(T4) and triiodothyronine (T3) when stimulated by TSH. Both are considered amine hormones, for each consists of a linkage of two iodinated tyrosine residues (Fig. 12-6). The 3 and 4 refer to the number of iodine atoms in their molecules. These hormones are necessary for normal growth and development in young animals, and they regulate basal metabolic rate in the adult.
Thyroid Gland. The thyroid gland is associated with the proximal part of the trachea near the thyroid cartilage of the larynx. its appearance varies widely among species, with the thyroid gland of most animals possessing two distinct lobes, variably connected across the midline by a strip of thyroid tissue called the isthmus. in pigs, the bulk of the gland lies primarily on the ventral aspect of the trachea rather than being clearly divided into lateral lobes. A connective tissue capsule covers the gland and gives rise to septa that divide the substance of the thyroid and support the vasculature of the gland. Arterial blood supply to the thyroid and the associated parathyroid glands (discussed later) arrives as branches of the common carotid artery.
Microscopically, the thyroid gland consists of follicles, spheres lined by a simple epithelium of cells that ranges from cuboidal to columnar (Fig.
12-7). Thyroid follicles are filled with the product of the follicular lining cells, a gel-like substance called colloid, which consists of a protein-iodine complex, thyroglobulin.Figure 12-7. Thyroid follicles of varying sizes and shapes are filled with colloid and lined with follicular cells. (Reprinted with permission of Wiley-Blackwell from Dellmann H.D. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
The hormones T3 and T4 are stored in the colloid as iodinated tyrosine residues that are part of thyroglobulin molecules (Fig. 12-8). This type of storage is a unique among endocrine glands.
scattered among the follicular lining cells and adjacent to them are a small subset of thyroid cells, the C cells (parafollicular cells). The C cells produce calcitonin, a peptide hormone that lowers the blood level of calcium by inhibiting the action of osteoclasts. Calcitonin release is directly regulated by the negative feedback of serum calcium concentration on C cells, not by TsH. The physiologic importance of calcitonin in the overall regulation of serum calcium concentration is minimal compared with the role of parathyroid hormone (discussed later).
Thyroxine and Triiodothyronine. Both T4 and T3 are biologically active (bind to thyroid hormone receptors), and most of the circulating hormones are bound to plasma proteins. The T4 and T3 in the body fluids that are not protein bound are the biologically active molecules, but they are also subject to degradation. in many species, the plasma levels of T4 are
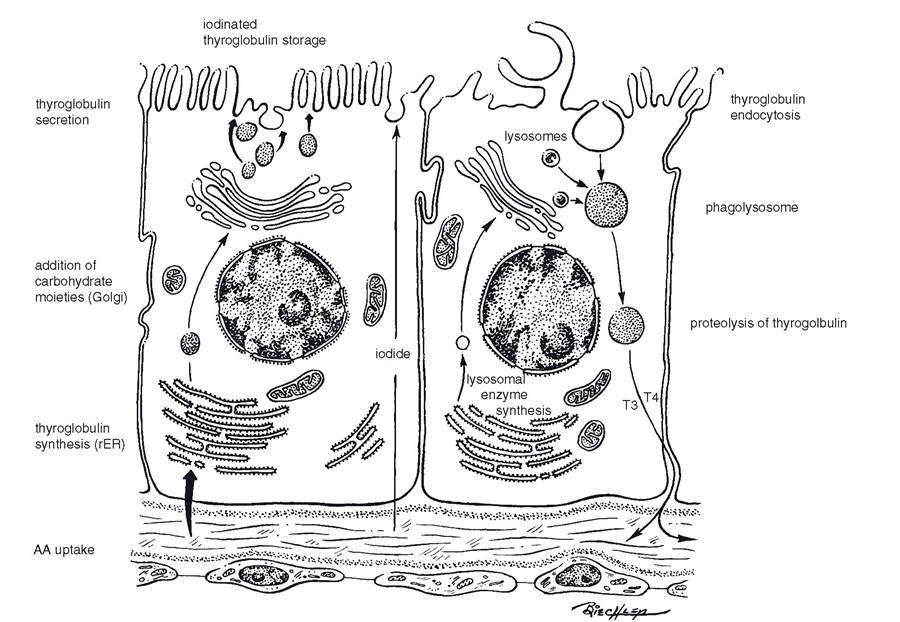
Figure 12-8. Biosynthesis of thyroglobulin (left) and its resorption, proteolysis, and secretion (right). Events are depicted in two cells, but they do occur in the same cell. (Reprinted with permission of Wiley-Blackwell from Dellmann H.D. Textbook of Veterinary Histology. 5th ed. Baltimore: Lippincott Williams & Wilkins, 1998.)
much higher than those of T3 because the affinity of T4 for plasma proteins is greater than T3. The intracellular receptors for the thyroid hormones bind both T4 and T3, but they have a higher affinity for T3. Because of the intracellular receptor affinity for T3 and the potential for T4 to be converted to T3 after it enters target cells, many consider T3 to be the more biologically important of the two hormones.
Secretion of T4 and T3 from the thyroid is a complex series of events that begins with the phagocytosis of thyroglobulin by follicular cells. Endocytotic vesicles containing thyroglobulin then fuse with lysosomes that contain enzymes necessary to degrade the thyroglobulin and release free T4 and T3 from their storage form as part of thyroglobulin molecules. The free T4 and T3 are then secreted into the blood (Fig. 12-8). TSH from the adenohypophysis stimulates follicular cells to synthesis thyroglobulin and to secrete T4 and T3 into the blood. Thus, the overall effect of TSH is to increase blood levels of the thyroid hormones.
Plasma levels of thyroid hormones are relatively stable in adult animals; unlike many other hormones, they do not show a significant diurnal rhythm. These stable levels are primarily maintained by negative feedback of T4 and T3 at the level of the pituitary. The thyroid hormones have direct effects on cells in the adenohypophysis to inhibit TSH synthesis and release. Thyrotrophin-releasing hormone (TRH) from the hypothalamus is always present and acts to promote TSH synthesis and release, but its levels do not respond to feedback regulation by T4 and T3.
Almost all tissues of the body are target tissues for thyroid hormones, for almost all have receptors for them. In mature animals the most general effect of thyroid hormones is to increase overall oxygen consumption and heat production. The basal metabolic rate is a measure of oxygen use in resting conditions, so thyroid hormones are said to increase it. The exact intracellular mechanisms responsible for these general effects are not known, but the effects are known as the calorigenic action of the thyroid hormones. The calorigenic action is associated with an overall increase in the metabolism and use of carbohydrates and lipids, which is consistent with the increased use of oxygen and heat production.
Chronic exposure of some animals to cold is associated with an increase in TSH, T4, and T3. The increased calorigenic effect of the thyroid hormones should work to maintain normal body temperature in the cold environment. The response to the cold appears to be a result of an increase in TRH release by the hypothalamus. The hypothalamus is known to be the reflex center for other reflexes involved with the regulation of body temperature on a short-term basis (e.g., peripheral vasodilation or vasoconstriction).
Thyroid hormones are essential for normal growth and development in young animals. Two systems of special importance are the skeletal and nervous systems. Animals with thyroid hormone deficits (hypothyroidism) do not attain normal stature and have a variety of central nervous system abnormalities. in humans with severe deficits at birth, mental development remains impaired throughout life, even if replacement therapy is begun when the person is 4 to 5 years of age. Cretinism is the term for the human condition caused by a congenital lack of thyroid hormone and characterized by arrested physical and mental development and lowered metabolic rate.