MODIFICATION OF GONADOTROPIN RELEASE
The main secretory pattern of gonadotropins is pulsatile-, the pattern is driven by pulsatile secretion of GnRH from the hypothalamus (Figure 35-4). The importance of this mode of delivery is shown by the fact that if GnRH is administered in a continuous (pharmacological) manner, the system can be downregulated.
Continual occupancy of GnRH receptors on gonadotrophs by GnRH interrupts the intracellular signal for the synthesis and release of gonadotropins. Successful induction of a fertile estrus in bitches can be performed by administering canine analogue GnRH; however, the dose must be diminished as ovulation approaches, or downregulation will occur.In general, the pulse generator system for gonadotropin secretion is increased in the follicular phase and decreased in the luteal phase of the estrous cycle (Figure 35-5). Estrogen decreases the pulse amplitude, and progesterone decreases the pulse frequency of gonadotropin secretion. This means that during the follicular phase, pulse frequency increases because of the absence of progesterone, and pulse amplitude decreases
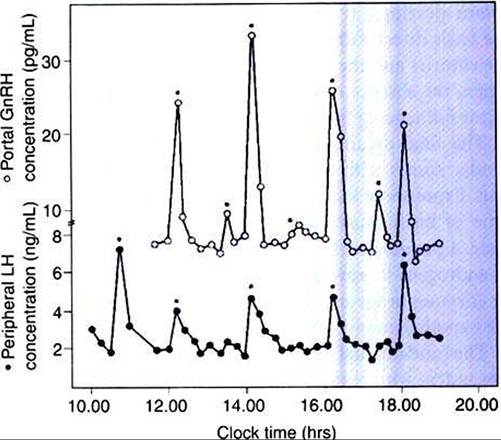
FIGURE 35-4 Concentrations of gonadotropin-releasing hormone (GnRH) in portal plasma (open circles) and luteinizing hormone (LH) in jugular venous plasma (solid circles) of four Ovariectomized ewes. Asterisks indicate secretory episodes (pulses) of GnRH and LH. (From Johnson M, Everitt B, editors: Essential reproduction, ed 3, London, 1988, Blackwell Scientific.)
because of the presence of estrogen. This combination of increased pulse frequency and decreased pulse amplitude is important for nurturing the final growth phase of the developing antral follicle.
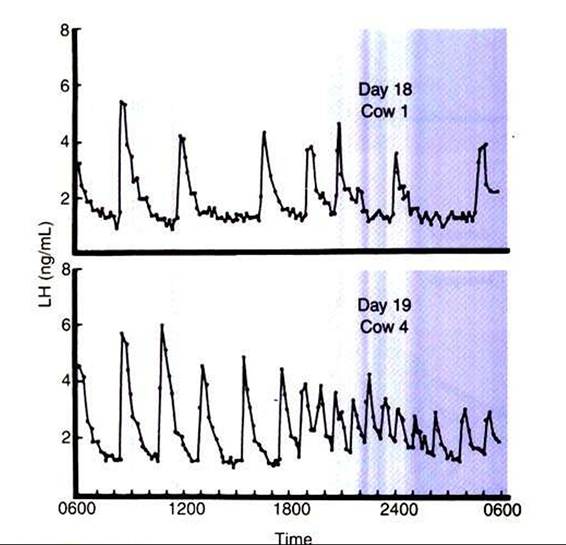
FIGURE 35-5 Pattern of plasma luteinizing hormone (LH) concentration on day 18 or 19 of the estrous cycle in two cows.
(From Rahe CH, Owens RE, Fleeger JL, et al: Pattern of plasma luteinizing hormone in the cyclic cow: dependence upon the period of the cycle, Endocrinology 107:498, 1980.)The hypothalamus and adenohypophysis are capable of responding to a sustained increase in estrogen secretion by increased secretion of gonadotropins, a relationship that is termed positive feedback. The sudden sustained increase in estrogen levels, which occurs over one to several days during final antral follicle development, causes an increase in gonadotropin secretion by increasing the frequency of pulsatile release of GnRH and, as a result, gonadotropin secretion. In essence, the frequency of pulsatile release of gonadotropins overcomes the metabolic clearance rate. The purpose of the gonadotropin surge is to induce changes within the follicle that lead to its rupture (ovulation). The duration of the gonadotropin surge is relatively short, usually 12 to 24 hours, possibly because the main factor driving the response, estrogen, declines in concentration as the follicles respond to the preovulatory gonadotropin surge. This particular physiological mechanism for initiating the onset of ovulation is effective because the follicle is able to signal its stage of maturity to the hypothalamus and adenohypophysis by a product (estrogen) that is produced in increasing amounts with increasing follicle maturity.
The secretion of gonadotropins is modified by the ovarian steroid hormones estrogen and progesterone. With time, the effect of these hormones is suppressive for gonadotropin secretion. Estrogens, in particular, cause negative-feedback inhibition of gonadotropin secretion, which is characterized by its sensitivity (effective at low concentrations) and its rapid onset (within a few hours). The substantial increase in gonadotropin concentrations that occurs after ovariectomy is caused largely by the removal of estrogens.
Because progesterone affects gonadotropin pulse frequency, it is thought that its modulatory effect is at the level of the hypothalamus.
Estrogens are thought to affect gonadotropin secretion through an effect on both the pituitary gland and the hypothalamus. Although there are differences in the site of action among species, it appears that the hypothalamic site for negative-feedback inhibition of gonadotropins by both progesterone and estrogen is in an area immediately above the median eminence, known as the arcuate nucleus. The hypothalamic site for positive-feedback stimulation of gonadotropin release by estrogen is probably further anterior, that is, in the preoptic anterior hypothalamic region.The secretion of gonadotropins can be modified by peptide and protein hormones produced by both the hypothalamus and the ovary. β-Endorphin, an opioid peptide produced from the hypothalamic precursor molecule pro-opiomelanocortin, can inhibit LH secretion when pharmacologically administered systemically. Its role in the physiological modulation of gonadotropin secretion, however, remains to be identified. Another hormone, inhibin, a protein produced by the granulosa cells of the developing follicle, also inhibits gonadotropin secretion, particularly FSH, during the final stages of follicle development. As described in the section on folliculogenesis, this depression of FSH secretion may be important to the animal for controlling the number of follicles that are brought to final maturation.
Control of gonadotropin secretion in the male is similar to that in the female; pulses of GnRH, arising in the hypothalamus, affect pulsatile secretion of the gonadotropins. This in turn causes the secretion of testosterone, also in pulsatile form, from the testes. One major difference between the genders is that the need for positive-feedback release of gonadotropins in males does not exist; gametes are produced and released on a continuous basis within a tubular system that opens to the exterior. This negates any need for a surge release of gonadotropins, as is required in the female to rupture the ovarian surface for the release of oocytes.
Prolactin is the third adenohypophysis-produced hormone that is important in the reproductive process, mainly because of its effect on the mammary gland and lactation in mammals. Although the secretion of prolactin is pulsatile, the control of secretion has more emphasis on inhibition than does stimulation of secretion. This concept is supported by the finding that prolactin secretion increases if the pituitary gland is disconnected from the hypothalamus by either cutting the pituitary stalk or transplanting the pituitary gland to another site (e.g., kidney capsule). Thus, most attention has focused on factors that inhibit prolactin secretion. The catecholamine dopamine, which is produced by neurons in the ventral hypothalamus (arcuate nucleus), is a potent inhibitor Ofprolactin secretion (Figure 35-6). Other factors that inhibit prolactin secretion are γ-aminobutyric acid (GABA) and GAP. Dopamine agonists, such as the ergot-type compounds bromocriptine and Cabergoline, can be used to suppress prolactin
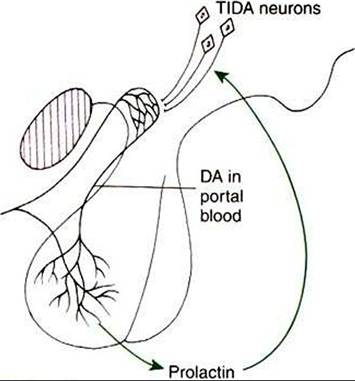
FIGURE 35-6 Diagrammatic summary of the proposed negative-feedback relationship between prolactin and dopamine (DA). Prolactin is believed to accelerate dopamine turnover in the arcuate nucleus neurons Ituberoinfundibular dopamine [TIDA] neurons), and the amine is then released into the portal capillaries to gain access to the Iactotropes. Hyperprolactinemia could be caused by either a failure of prolactin inhibitory factor activity at the dopamine receptor level in the anterior pituitary or a reduction ofTIDA neuron activity in the hypothalamus. (From Johnson M, Everitt B, editors: Essential reproduction, ed 3, London, 1988, Blackwell Scientific.)
secretion in cases of hyperprolactinemia. Cabergoline, a potent prolactin inhibitor, can be used to shorten interestrous intervals in female dogs and promote Iuteolysis in female dogs and female cats during the latter half of pregnancy (prolactin is a Iuteotropin). The negative-feedback control of prolactin is shown in Figure 35-6.
One of the first known prolactin-releasing factors was thyrotropin-releasing hormone (TRH). The physiological relevance of TRH in prolactin secretion is still unknown despite that receptors for TRH have been identified on lactotropes within the adenohypophysis. Vasoactive intestinal peptide (VIP), a potent stimulator of prolactin secretion, may play a physiological role in prolactin secretion through inhibition of dopamine synthesis within the hypothalamus. Estrogens can increase prolactin secretion by Iactotropes by decreasing Iactotrope sensitivity to dopamine and increasing the number of TRH receptors. Interestingly, female dogs undergoing ovariohysterectomy with a cesarean section usually maintain the ability to lactate effectively afterward.