NERVOUS SYSTEM
The avian brain is small with less development of the cerebral hemispheres. These are composed mainly of the corpora striata, indicating that birds use less learning and memory but more instinct and stereotypical behavior.
More intelligent birds like parrots have better development of the cerebral hemispheres.There is a well-developed cerebellum for locomotion and large optic lobes for vision. There are 12 pairs of cranial nerves, as in mammals. The spinal cord resembles that of mammals, having three meninges: the dura, arachnoid, and pia mater. However, as the cord is the same length as the canal there is no cauda equina (King & McLelland 1984). The spinal cord becomes enlarged at the brachial and lumbosacral plexi. Flying birds have a more prominent brachial plexus, while running birds like the ostrich have a large lumbar plexus (Rosenthal 1997a). A unique feature of birds is the “glycogen body.” This pea shaped, glycogen-rich cleft lies on the dorsal surface of the lumbosacral plexus but its function is unknown (Rosenthal 1997a).
The roots of the lumbar plexus are in contact with the dorsal surface of the cranial kidney; the sacral plexus is totally embedded within its middle division. Some of the roots of the pudendal plexus are embedded in the caudal division.
Senses
The fact that birds fly mean they must have good aerial vision, but olfaction is less essential due to the absence of odors in the sky. Hence, in the avian brain the olfactory bulbs are reduced while the optic lobes are increased. Recognition of food relies mainly on sight and, to a lesser degree, by feel (ducks and geese) and smell.
Sight
The large eyes and well-developed optic lobes in the brains means birds have excellent vision. In fact, in many species the two eyes weigh more than the brain (Evans 1996; Gunturkun 2000). Most birds also have a high degree of binocular and color vision.
The optic nerve is the most developed cranial nerve in birds, being largest in Corvids and Falconiformes and smallest in nocturnal species. There is no consensual light reflex in birds because there is complete decussation of the optic nerve fibers at the optic chiasma (King & McLelland 1984).External adnexa
Although the eyelids are open and well developed in precocial birds, they are sealed in altricial birds. Eyes usually open between 2 and 4 weeks and usually take a few days for full separation.
Birds have upper and lower eyelids and a nictitating membrane. The lower lid is far more mobile and covers more of the eye while blinking than the upper (Martin 1985). Modified feathers, called filoplumes, are on the lids and act like mammalian cilia.
The lids close mainly in sleep. Blinking is performed mainly by the mobile nictitating membrane (30-35 times per minute in domestic fowl). This is usually transparent in diurnal species so that vision is not impaired.
Orbital glands
The Harderian gland is the largest. It lies craniomedial within the orbit behind the nictitating membrane and produces a mucoid secretion that moistens the cornea. The lacrimal gland lies at the caudolateral margins. Both glands empty via dorsal and ventral puncta into the nasolacrimal duct. The nasal (salt) gland lies dorsomedial to the orbit and empties separately into the nasal cavity (Martin 1985).
Globe
The eyeball consists of a small, unprotected anterior portion covered by the cornea, and the posterior part that is protected by the two orbits separated by a thin bony septum. The anterior part has three basic shapes: flat, globose or tubular, depending on the species (Figs. 6.65 and 6.66, Table 6.5). The shape of the eyeball is formed by 10 to 18 scleral ossicles, which are visible by radiography (Fig. 6.67). These are a ring of overlapping bones, which strengthen the eye and provide an attachment for the ciliary muscles, permitting greater accommodation (Martin 1985).
They are largest in owls and diurnal raptors and less developed in Psittaciformes and Anseriformes.The eyeball fills the orbit so the extraocular muscles are less developed than in mammals (Kern 1997; Martin 1985). The single occipital neck joint and long, flexible neck compensate instead by allowing the bird to rotate its neck. Movements of each eye are independent of each other.
Iris
This is usually dark in color, ranging from brown to black, but in owls it can be bright yellow and it is pale blue in gannets. It can also vary with diet as it contains lipid droplets. In some species it can help identify age; for example, the African Gray parrot has a gray iris when juvenile that becomes yellow when mature. In contrast to mammals, the muscles are striated, making them under some voluntary control, so atropine will be ineffective (Fig. 6.68). Dilation of the avian iris is only effective with mydriatics such as the muscle relaxant vecuronium.
CLINICAL NOTE
The pupil is round and, as the iris has striated muscle, it cannot be dilated with normal mydriatics. There is no consensual light reflex (Fig. 6.68).
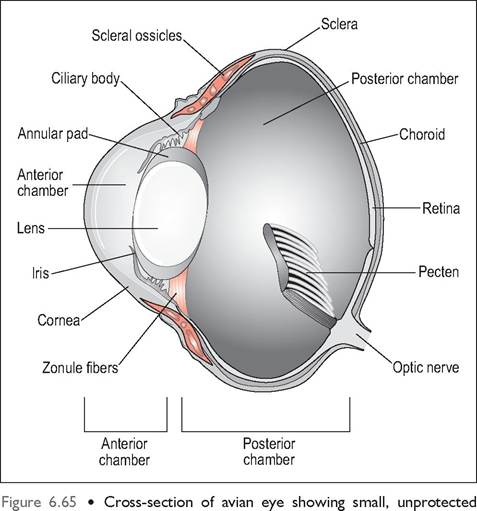
anterior portion covered by the cornea and large posterior part with pecten. The intermediate area is supported by a ring of scleral ossicles.
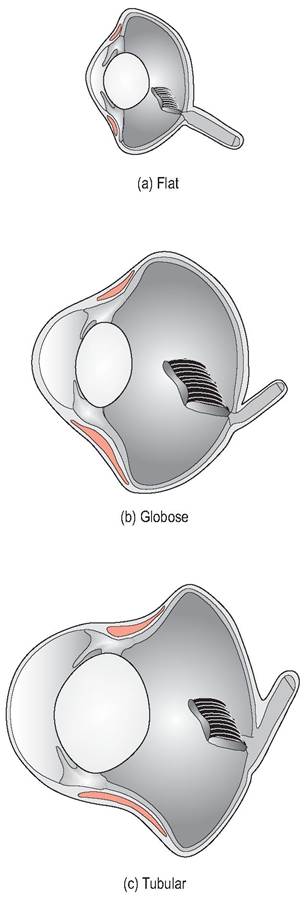
Figure 6.66 • The anterior part of the avian eye has three basic shapes depending on species.
(a) Flat
(b) Globose
(c) Tubular
| Table 6.5 Basic avian globe shapes (King & McLelland 1984) | ||
| Globe shape | Species | Vision |
| Flat Globose Tubular | Most birds e.g., Chicken Birds of prey, Passerines Nocturnal birds of prey | Poor visual acuity Good visual acuity Excellent visual acuity |
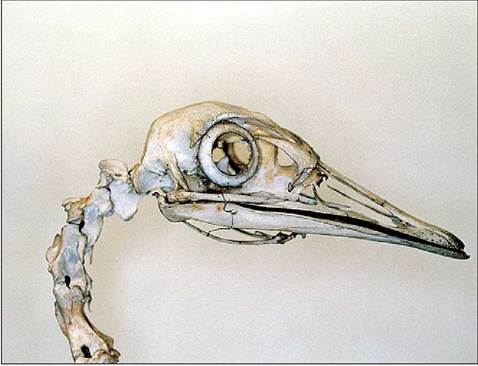
Figure 6.67 • Skull of rhea (Rhea americana) showing ring of scleral ossicles.

Figure 6.68 • Birds have round pupils and the iris has striated muscles which cannot be dilated with normal mydriatics like atropine.
Lens
Accommodation is very well developed in birds, with a softer lens to allow for rapid accommodation. There are three mechanisms for this. Diurnal birds use the posterior sclerocorneal muscles to compress the lens while nocturnal birds and hawks use the anterior sclerocorneal muscle. In diving birds the cornea has poor accommodation underwater so they use the combined action of sclerocorneal muscles and the iris sphincter muscle.
Retina
In order to attain high visual acuity, the avian retina lacks blood vessels and tapetum lucidum to prevent shadows and scattering of light. Instead, it is vascularized by a black, vascular comb-like structure called the pecten. This structure extends from the optic disk into the vitreous body towards the lens and is unique to birds. It appears to nourish the relatively avascular retina, aid in acid-base balance, and facilitate fluid movement within the eye (Gunturkun 2000; Martin 1985).
The fundus is usually gray or reddish. The optic disk is elongated and oval but is mainly obscured by the pecten. Cones, which provide visual acuity and excellent color vision, are more numerous in diurnal species, forming up to 80% in the chicken and pigeon. Nocturnal birds have more rods than cones.
CLINICAL NOTE
Many diurnal birds lack rods so have excellent day vision and poor night vision, which is why dimming the lights makes it easier to catch birds (King-Smith 1971).
Fovea
The fovea is the area of densely packed cone cells and, in birds, is deep for greater visual acuity. Most birds have a central fovea but ground dwellers have none, and Falconi- formes, which have the celebrated “eagle eye”, have two to allow high visual acuity (Kern 1997; Martin 1985).
Hearing
After vision, hearing is the second most important avian sense.
Birds, especially diurnal ones, generally have the same hearing ability as humans. However, nocturnal birds like the owl have an amazingly accurate ability to pinpoint sound, and homing pigeons can hear extremely low frequencies. Some nocturnal and underwater species use echoes to locate obstacles while flying or diving. Birds also have the ability to hear sounds of rapid frequency because the syrinx of songbirds is capable of producing a duet of high pitch inaudible to the human ear.Like mammals, the ear is the organ of hearing and balance and is divided into the external, middle and inner ear.
External ear
There is no pinna and the external ear opening is hidden by feathers called ear coverts. The surrounding skin is loose and can be drawn forward by the dermo-osseus muscle to reduce the opening to a vertical slit. This flap can also close over to block sound entering the ear, in for example, the crowing cock (King-Smith 1971). In some species, such as owls, a vertical skin flap called the operculum lies rostral to the external ear opening (Fig. 6.70). This has a row of stiff feathers at right angles and this facial ruff can be erected by skeletal muscle to help locate sound, equivalent to someone cupping their hands behind their ears. Acute hearing is also facilitated in some owl species by asymmetrical ear positioning, allowing one ear to hear sounds on a horizontal plane and the other on a vertical plane (King & McLelland 1984; Necker 2000).
Middle ear
The middle ear is the air-filled cavity between the tympanic membrane and inner ear. There is only one bony ossicle called the columella (equivalent of the mammalian stapes) which transmits sound vibrations from the tympanum to
Figure 6.70 • Long eared owl (Asio otus) showing the operculum. This is a flap of feathered skin rostral to the external ear opening which can be raised to capture sound on the same principle as a satellite dish.
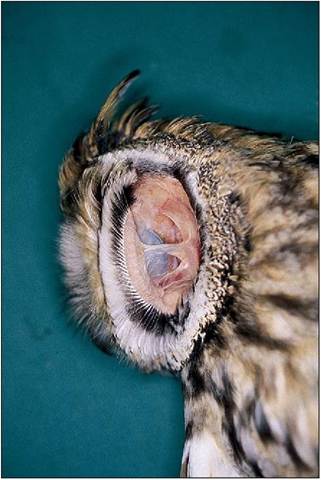
the vestibular window of the inner ear (Necker 2000). The middle ear is connected to the oropharynx by the pharyngotympanic tubes (the equivalent of mammalian eustachian tubes). As birds need to adjust rapidly to high altitudes, these tubes are short and open directly into the pharynx via the common infundibular cleft (Evans 1996; King & McLelland 1984) (Fig. 6.69).
Inner ear
This contains the organ of hearing (the cochlea) and the organs of balance (the three semicircular canals, utricle, and saccule). Unlike in mammals, the cochlea is short and not coiled and the semicircular canals are larger and more thick walled. The eighth cranial nerve receives both vestibular and auditory sensory fibers.
Olfaction
The olfactory system consists of the external nares and the caudal conchae that are lined by olfactory epithelium and connected to the olfactory bulbs of the brain. There is no vomeronasal organ (Mason & Clark 2000).
Birds do have a sense of smell, although it may not be as highly developed as vision and hearing (King-Smith 1971). Many species use olfactory cues to find food, and smell plays a role in maternal identification of young (Mason & Clark 2000). The olfactory bulbs are smallest in seed eaters and larger in aquatic birds. The ground-dwelling nocturnal kiwi, which eats earthworms, has the largest of all and is the only bird with nostrils at the end of its beak. Vultures, which feed on dead carrion, also have a well-developed sense of smell, and homing pigeons use olfactory cues to find their way home (King & McLelland 1984).
Taste
The taste buds are confined to glandular non-cornified epithelium. They lie at the base of the tongue, and in the roof and floor of the oropharynx. In parrots they lie at either side of the choana and at the rostral end of the laryngeal mound.
The total number of taste buds is far fewer than in mammals, with the chicken having 24 taste buds and parrots 350, in contrast to 9000 in man and 17,000 in the rabbit (Klasing 1998; Mason & Clark 2000). Birds, however, do respond to salts and acids and dislike bitter-tasting substances. They are also far more tolerant of spicy foods than mammals. For example chili red peppers (capsaicin), which would be irritating to mammals, have little effect on birds. Parrots and humming birds have a definite preference for sugary water, while birds with salt glands have a higher preference for salt tastes (King & McLelland 1984; Mason & Clark 2000).
Touch
Birds have widely distributed mechanoreceptors called Herbst corpuscles in their skin. These are located deep in the dermis and found on beaks, legs and associated with feather follicles. In the plumage these are used to ruffle feathers and help stimulate preening.
Avian species that use their beak to search and locate food also have these receptors located in a well-developed “beak tip organ”, which is located along the edge and tip of the upper beak in aquatic birds like ducks and geese (Gerlach 1997b). Psittacines have them located in the lower beak. These remarkably sensitive touch receptors compensate for the lack of taste buds. For example, mallard ducks have more sensation in their beak tip than we humans have in our index finger tip (Klasing 1998).
KEY POINTS
• Birds have excellent hearing and sight but poor sense of smell and taste.
• There is no external ear and only one middle ear bone, called the columella.
• The eyes have scleral ossicles. An avascular retina is nourished by the pecten, and fovea are present for visual acuity.
• There is no avian consensual light reflex.
• The iris is controlled by striated muscle pupil and so is non responsive to atropine.
• Neck rotation compensates for poor orbital muscles. Eyes can, however, move independently.